Article Text

Abstract
If a patient asks ‘why does my shoulder hurt?’ the conversation will quickly turn to scientific theory and sometimes unsubstantiated conjecture. Frequently, the clinician becomes aware of the limits of the scientific basis of their explanation, demonstrating the incompleteness of our understanding of the nature of shoulder pain. This review takes a systematic approach to help answer fundamental questions relating to shoulder pain, with a view to providing insights into future research and novel methods for treating shoulder pain. We shall explore the roles of (1) the peripheral receptors, (2) peripheral pain processing or ‘nociception’, (3) the spinal cord, (4) the brain, (5) the location of receptors in the shoulder and (6) the neural anatomy of the shoulder. We also consider how these factors might contribute to the variability in the clinical presentation, the diagnosis and the treatment of shoulder pain. In this way we aim to provide an overview of the component parts of the peripheral pain detection system and central pain processing mechanisms in shoulder pain that interact to produce clinical pain.
- Molecular biochemistry
- Shoulder injuries
- Tendons
- Orthopaedics
- Neurology
Statistics from Altmetric.com
Introduction: a very brief history of pain science essential for clinicians
The nature of pain, in general, has been a subject of much controversy over the past century. In the 17th century Descartes’ theory1 proposed that the intensity of pain was directly related to the amount of associated tissue injury and that pain was processed in one distinct pathway. Many earlier theories relied upon this so-called ‘dualist’ Descartian philosophy, seeing pain as the consequence of the stimulation of a ‘specific’ peripheral pain receptor in the brain. In the 20th century a scientific battle between two opposing theories ensued, namely specificity theory and pattern theory. The Descartian ‘specificity theory’ saw pain as a specific separate modality of sensory input with its own apparatus, while ‘pattern theory’ felt that pain resulted from the intense stimulation of non-specific receptors.2 In 1965, Wall and Melzack's3 gate theory of pain provided evidence for a model in which pain perception was modulated by both sensory feedback and the central nervous system. Another huge advance in pain theory at around the same time saw the discovery of the specific mode of actions of the opioids.4 Subsequently, recent advances in neuroimaging and molecular medicine have vastly expanded our overall understanding of pain.
So how does this relate to shoulder pain? Shoulder pain is a common clinical problem, and a robust understanding of the way in which pain is processed by the body is essential to best diagnose and treat a patient's pain. Advances in our knowledge of pain processing promise to explain the mismatch between pathology and the perception of pain, they may also help us explain why certain patients fail to respond to certain treatments.
Basic building blocks of pain
Peripheral sensory receptors: the mechanoreceptor and the ‘nociceptor’
There are numerous types of peripheral sensory receptors present in the human musculoskeletal system.5 They may be classified based on their function (as mechanoreceptors, thermoreceptors or nociceptors) or morphology (free nerve endings or different types of encapsulated receptors).5 The different types of receptor can then be further subclassified based on the presence of certain chemical markers. There are significant overlaps between different functional classes of receptor, for example between the mechanoreceptor and nociceptor, and this has the potential to cause significant confusion. For this reason it is absolutely vital to understand that there is no universally accepted receptor classification system and that receptors are best seen as a continuum, in which the artificial boundaries between different receptor types are somewhat arbitrary and fluid. Despite this, it is still worthwhile considering how sensory receptors may be classified in greater detail.
Sensory receptors are supplied by afferent nerves of varying sizes, degrees of myelination and conduction velocities. On this basis sensory nerves can be simply divided into three main groups (conduction velocity given in brackets): thick diameter myelinated group II or Aβ fibres (>20 m/s), small diameter myelinated group III or Aδ fibres (2.5–20 m/s) and unmyelinated group IV or C fibres (<2.5 m/s).6 Receptors are sometimes referred to simply by the characteristics of the nerves that innervates them, for example ‘Aδ nerve endings’. This description makes no reference to a receptor's other specific characteristics as outlined above and described in greater detail in the following.
Receptors that respond preferentially to noxious stimuli and those that have a high threshold to the adequate stimulus are termed ‘nociceptors’.7 Nociceptors may respond to multiple energy forms such as thermal, mechanical and chemical stimuli. Nociceptors can be comprehensively subclassified based on four criteria: myelination of nerve supply (unmyelinated C fibres versus myelinated Aδ fibres), modalities of stimulus that evoke a response, response characteristics and distinctive chemical markers.8 On a molecular basis, nociceptors can be either peptidergic or non-peptidergic based on whether peptides are expressed in their dorsal root ganglion cells.8 The peptides expressed include substance P (SP), calcitonin gene-related peptide (CGRP) and somatostatin. The subclassification of nociceptors is complex; furthermore, they have a degree of functional and chemical plasticity which ensures that their threshold and responsiveness, as well as the efficacy of their synaptic contacts, are regulated to reflect changes produced by activity, inflammation and axonal injury.8
The second form of receptors is the mechanoreceptor which conveys information about the mechanical stimuli to the central nervous system. The main innocuous-touch receptors include (skin stimulus is in brackets): Meissner corpuscles (dynamic deformation), Merkel cell–neurite complexes (indentation depth), Pacinian/lamellar corpuscles (vibration), Ruffini receptors/endings/corpuscles (stretch) and free nerve endings (touch). The term ‘proprioceptor’ is used to describe a mechanoreceptor's role which is that of responding to the mechanical variables associated with muscles and joints.9 Proprioceptors include muscle spindles, Golgi tendon organs and Ruffini-type receptors.
Mechanoreceptors can be classified as low-threshold mechanoreceptors (LTMs), thus called because they respond to innocuous mechanical forces, or high-threshold mechanoreceptors (HTMs), thus called because they are excited only by greater mechanical forces. As with nociceptors, different types of mechanoreceptors are associated with specific afferent nerve types. Undoubtedly, there is a significant degree of crossover between the properties of both nociceptors and mechanoreceptors, demonstrated by the fact that HTMs can also be described as nociceptors.10 ,11 Certainly, creating any absolute division between the two is somewhat artificial and potentially misleading, especially given the plasticity of the receptor characteristics.
Generally, mechanoreceptors supplied by Aβ nerve endings are LTMs, while those supplied by Aδ nerve endings may be either HTMs or LTMs. HTMs supplied by Aδ nerve endings and the majority of C fibre nerve endings may be termed nociceptors. Schaible and Grubb6 have studied the mechanosensitivity of different fibre types (Aβ, Aδ and C) supplying the cat knee joint. They found that most Aβ nerve endings were activated by innocuous stimuli and hence could be termed LTMs. A much higher percentage of Aδ and C fibre nerve endings were insensitive to innocuous stimuli (50% and 70%, respectively) and could be classified as HTMs. Many LTMs fire in the innocuous range but have a far stronger response in the noxious range, demonstrating the dynamic complexity of their function.
A typical joint nerve is innervated by thick diameter myelinated Aβ, small diameter myelinated Aδ and a high proportion (circa 80%) of unmyelinated C fibres. C fibres are further subdivided into either sensory afferents or sympathetic efferents, with about 50% of each.6 Muscle nerves consist of axons from motor neurons, sensory neurons and postganglionic sympathetic neurons. Each of these nerve types may end in a number of different ‘receptors’ as described hereinbefore. Freeman and Wyke12 classified the different types of articular nerve endings based on their pioneering work in the knee joints of cats in 1967, these being summarised in table 1. They classified the different endings as types I, II, III and IV on a morphological and functional basis. Some of the eponymous terms used to describe receptors of different types are included in table 1, for example ‘Ruffini ending’ and ‘Golgi-Mazzoni body’. Other authors have since modified the eponymous terms mentioned in Freeman and Wyke's classification system.13
The Freeman and Wyke classification of joint nerve endings
Articular Aβ fibres terminate as corpuscular endings of the Ruffini, Golgi and Pacini types in the fibrous capsule, articular ligaments, menisci and adjacent periosteum.14 While articular Aδ and C fibres terminate as non-corpuscular or free nerve endings in the fibrous capsule, adipose tissue, ligaments, menisci, periosteum and synovium. The cartilage is not innervated. In muscles, thick myelinated afferents terminate as organised endings such as muscle spindles and tendon organs, while Aδ and C fibres terminate as free nerve endings. The receptor types and innervation specifically relating to the shoulder will be explored in greater detail in sections ‘Where are these receptors located in the shoulder?’ and ‘Neural anatomy of the shoulder’.
Peripheral pain processing: ‘nociception’
Tissue injury involves a variety of inflammatory mediators being released by damaged cells including bradykinin, histamine, 5-hydroxytryptamine, ATP, nitric oxide and certain ions (K+ and H+). The activation of the arachidonic acid pathway leads to the production of prostaglandins, thromboxanes and leukotrienes. Cytokines, including the interleukins and tumor necrosis factor α, and neurotrophins, such as nerve growth factor (NGF), are also released and are intimately involved in the facilitation of inflammation.15 Other substances such as excitatory amino acids (glutamate) and opioids (endothelin-1) have also been implicated in the acute inflammatory response.16 ,17 Some of these agents may directly activate nociceptors, while others bring about the recruitment of other cells which then release further facilitatory agents.18 This local process resulting in the increased responsiveness of nociceptive neurons to their normal input and/or the recruitment of a response to normally subthreshold inputs is termed ‘peripheral sensitisation’. Figure 1 summarises some of the key mechanisms involved.

Peripheral sensitisation and inflammation at the peripheral nerve terminal (ASIC, acid sensing ion channel; iGluR, inotropic glutamate receptor; IL-1β, interleukin-1β; IL-6, interleukin-6; mGluR, metabotropic glutamate receptor; NGF, nerve growth factor; PGE2, prostaglandin E2; PKA, protein kinase A; PKC, protein kinase C; TNF-α, tumour necrosis factor α; TrkA, tyrosine kinase receptor A; TTXr, tetrodotoxin-resistant sodium channel; μ, mu-opioid receptor; M2, muscarinic receptor; 5-HT3, serotonin; H1, histamine receptor; B1/B2, bradykinin receptors; A2, adenosine receptor; GABAR, γ-aminobutyric acid receptor; EP, prostaglandin receptor; VEGF, vascular endothelial growth factor). Access the article online to view this figure in colour.
NGF and the transient receptor potential cation channel subfamily V member 1 (TRPV1) receptor have a symbiotic relationship when it comes to inflammation and nociceptor sensitisation. The cytokines produced in inflamed tissue result in an increase in NGF production.19 NGF stimulates the release of histamine and serotonin (5-HT3) by mast cells, and also sensitises nociceptors, possibly altering the properties of Aδ fibres such that a greater proportion become nociceptive. The TRPV1 receptor is present in a subpopulation of primary afferent fibres and is activated by capsaicin, heat and protons. The TRPV1 receptor is synthesised in the cell body of the afferent fibre, and is transported to both the peripheral and central terminals, where it contributes to the sensitivity of nociceptive afferents. Inflammation results in NGF production peripherally which then binds to the tyrosine kinase receptor type 1 receptor on the nociceptor terminals, NGF is then transported to the cell body where it leads to an up regulation of TRPV1 transcription and consequently increased nociceptor sensitivity.19 ,20 NGF and other inflammatory mediators also sensitise TRPV1 through a diverse array of secondary messenger pathways. Many other receptors including cholinergic receptors, γ-aminobutyric acid (GABA) receptors and somatostatin receptors are also thought to be involved in peripheral nociceptor sensitivity.
A large number of inflammatory mediators have been specifically implicated in shoulder pain and rotator cuff disease.21–25 While some chemical mediators directly activate nociceptors, most lead to changes in the sensory neuron itself rather than directly activating it. These changes may be early post-translational or delayed transcription dependent. Examples of the former are changes in the TRPV1 receptor or in voltage-gated ion channels resulting from the phosphorylation of membrane-bound proteins. Examples of the latter include the NGF-induced increase in TRV1 channel production and the calcium-induced activation of intracellular transcription factors.
Primary hyperalgesia and peripheral sensitisation
Primary hyperalgesia is defined as hyperalgesia, or hypersensitivity, at the site of injury (the primary zone).26 Several mediators may lead to the sensitisation of a nociceptor and therefore play a role in primary hyperalgesia.27 Primary hyperalgesia relates to both heat and mechanical stimuli.28 The hyperalgesia to heat stimuli that occurs at the site of injury is thought to be owing to the sensitisation of primary afferent nociceptors, that is, peripheral sensitisation. However, the hyperalgesia to mechanical stimuli at the site of injury results from not only peripheral sensitisation, but probably from central sensitisation as well.29 Different types of mechanical hyperalgesia have been described: one type is hyperalgesia to light touch or ‘allodynia’, another is ‘punctate’ hyperalgesia and a third is termed ‘pressure’ or ‘impact’ hyperalgesia.30 It must be remembered that the acute presence of both allodynia and ‘punctate’ hyperalgesia at the site of injury may be demonstrative of a completely normal pain response, in response to acute injury for example, sunburnt skin. A lowered threshold to stimulation, an increased response to suprathreshold stimuli and the expansion of receptive fields are all ways in which the nociceptor sensitisation of primary hyperalgesia may be manifest.31
Secondary hyperalgesia and central sensitisation
Secondary hyperalgesia is defined as hyperalgesia outside the original zone of injury and relates to the sensory response to mechanical stimuli only.28 The localised activation of nociceptors leads to a ‘flare response’ outside the zone of initial injury through the spreading chemical activation of adjacent nociceptors. However, La Motte et al32 demonstrated that proximal nerve blocks prevent the development of secondary hyperalgesia, indicating that higher pathways must be involved and that the flare response cannot fully account for secondary hyperalgesia. The flare response and secondary hyperalgesia have also been shown to be distinct entities for several reasons: the zone of secondary hyperalgesia is generally larger than the ‘flare’, they can exist independently, and secondary hyperalgesia does not migrate across the body's midline, whereas flare does.32 Peripheral sensitisation cannot adequately account for secondary hyperalgesia; hence the central nervous system must have a key role to play in this phenomenon.27 ,33 It is clear that the peripheral signal for pain is not purely driven by nociceptors; under pathological circumstances other receptors types, which are normally associated with the sensation of touch, acquire the ability to elicit pain.33 ,34 This applies to neuropathic pain as well as to acute secondary hyperalgesia.
The term ‘central sensitisation’ is used to describe the phenomenon whereby central pain-signalling neurons become hypersensitive to the input of LTMs.34 ‘Central sensitisation’ may also be more generally described as ‘an amplification of neural signalling within the CNS that elicits pain hypersensitivity’.35 Secondary hyperalgesia (and therefore central sensitisation) is seen in many clinical situations including the acutely inflamed joint,36 the arthritic knee37 and the painful shoulder.38
Two distinct forms of secondary hyperalgesia have been observed: punctuate and light touch hyperalgesia (allodynia). Punctate hyperalgesia is thought to be different from light touch hyperalgesia for several reasons: the area of punctuate hyperalgesia is larger than that of light touch hyperalgesia and light touch hyperalgesia post capsaicin is significantly shorter lived than punctate hyperalgesia. Light touch hyperalgesia32 and punctate hyperalgesia33 both appear to be mediated by LTMs. Ziegler et al39 used an intradermal capsaicin model to show that punctuate hyperalgesia was mediated by smaller myelinated fibres than light touch hyperalgesia. Certainly, the central sensitisation is induced by unmyelinated C fibre nociceptors, but the exact higher mechanisms have yet to be fully determined40: it is likely that the sensitisation of nociceptive neurons in the dorsal horn of the spinal cord are involved, although a novel presynaptic model has been proposed by Cervero.41 The way in which the facilitated pathways (myelinated A fibres) are separate from the facilitating pathways (unmyelinated C fibres) in this process is called a ‘heterosynaptic’ form of facilitation.29 Heterosynaptic facilitation is distinct to ‘homosynaptic’ facilitation, an example of the latter being ‘wind up’, that is, the temporal summation to punctuate stimuli in the zone of capsaicin injection. In ‘homosynaptic’ facilitation, the events responsible for triggering synaptic strengthening occur at the same synapse that is being strengthened. In ‘heterosynaptic’ facilitation synaptic strengthening between a presynaptic and a postsynaptic cell may occur as a result of the firing of a third neuron, a modulatory interneuron, whose terminals end on and regulate the strength of the specific synapse.42
Spinal cord
The spinal dorsal horn receives inputs from a wide variety of primary afferent fibres including those from nociceptors and mechanoreceptors.40 Primary afferents generally use glutamate as their principal neurotransmitter and form excitatory (glutaminergic) synapses with the neurons in the dorsal horn.40 The primary afferent fibres may synapse with two broad types of neuron: projection cells which travel in rostral parts of the spinal cord to the higher brain centres and interneurons which remain in the spinal cord and contribute to local neuronal circuits. Interneurons can be either inhibitory, using GABA/glycine as their neurotransmitter, or excitatory (glutaminergic).43 The intrathecal administration of drugs that antagonise GABA/glycine receptors can cause allodynia, suggesting that one key function of inhibitory interneurons is to suppress the activity of tactile afferents so that they do not normally elicit pain. The dorsal horn also receives an important descending input from higher brain centres, these systems are serotonergic (5-HT) and norepinephrinergic, with 5-HT3 receptors and α2 adrenoceptors being found in the superficial dorsal horn.43 Stimulation of these descending inhibitory circuits produces analgesia; the mechanism behind this appears to be related to inhibitory actions on excitatory interneurons and projection neurons. These descending systems may be called the descending pain modulatory system (DPMS).
Examples of the synaptic plasticity of the dorsal horn include the activity-dependent plasticity of ‘wind-up’, the ‘heterosynaptic’ central sensitisation as described earlier, the 'homosynaptic’ increases in response seen in long-term potentiation and transcription-dependent central sensitisation.15 Wind-up's summative increase in dorsal horn neuron output is related to the SP and glutamate-induced change in dorsal horn postsynaptic sensitivity.44 Heterosynaptic central sensitisation is partly related to transmitters and mediators released from primary afferents and glial cells, these then act at a distance on dorsal horn neurons to produce long-lasting heterosynaptic potentiation of fast synaptic transmission: the two general mechanisms implicated in this are the post-translational processing of ion channels/receptors/regulatory proteins and the cell surface expression/trafficking of ion channels.34 Long-term potentiation is the homosynaptic increase in response to primary afferent inputs and NMDA/AMPA receptor-activated pathways.45 ,46 Transcription-dependent central sensitisation involves multiple signalling pathways in the sensory and dorsal horn neurons.45 Figure 2 summarises our current understanding of some of the mechanisms behind central sensitisation in the spinal cord.

Mechanism underlying central sensitisation in the spinal cord (AMPAR, AMPA receptor; NMDAR, NMDA receptor; KaiR, kainite receptor; P2X, purinoreceptor; Src, Src tyrosine kinase; MAPK, mitogen-activated protein kinase; PKC, protein kinase C; NK1R, neurokinin-1 receptor; mGluR, metabotropic glutamate receptor; SP, substance P; Glu, glutamate; P, phosphate residue; VGCC, voltage-gated calcium channel). Access the article online to view this figure in colour.
Brain
Nociceptive information is transmitted from the spinal cord to the brain via several different pathways.15 These ascending pathways include direct projections to the thalamus (spinothalamic tract), direct projections to homeostatic control regions (spinomedullary and spinobulbar) and projections to the hypothalamus/ventral forebrain (spinohypothalamic). The periaqueductal grey (PAG) and reticular formation are important parts of the spinobulbar system. The thalamus is a key structure in pain processing and has ascending projections to multiple areas including the primary and secondary somatosensory cortices, the anterior insular cortex and the cingulate cortex.47 The subjective experience of pain is highly complex and involves other regions such as the amygdala, prefrontal cortex, cerebellum and basal ganglia. Key modulatory circuits involving the rostral ventromedial medulla (RVM) and PAG exert bidirectional control over dorsal horn nociceptive transmission.47 This network receives numerous direct and indirect inputs from pain pathways that include the amygdala, the anterior cingulate cortex and the anterior insula; providing a mechanism for the way in which emotion may affect pain perception. The RVM has separate descending antinociceptive and nociceptive outputs to the dorsal horn. Figure 3 summarises some of the key pain modulating circuitry.

Pain modulating systems (PAG, periaqueductal grey, RVM, rostroventromedial medulla, DLPT, dorsolateral pontine tegmentum). Access the article online to view this figure in colour.
Where are these receptors located in the shoulder?
The peripheral receptors play a pivotal role in the way in which tissue pathology may generate clinical pain. Therefore, understanding both the location of the receptors and the types of receptor present in the shoulder are of key importance in diagnosing and treating shoulder pain.
Rotator cuff muscles and tendons
Minaki et al48 investigated the innervation of the rabbit rotator cuff electrophysiologically and demonstrated a high density of nociceptors to be present around the rotator cuff's humeral insertion. Supraspinatus was particularly densely innervated with both mechanoreceptors and nociceptors as shown in figure 4 which shows the locations of receptive fields identified in the posterior shoulder. In a histological study in rats Backenkohler et al49 showed that Golgi tendon organs were found within tendons where they merge into muscle and in the tendinous insertions near the joint capsule; it was also shown that the muscle spindles tended to accumulate near the musculotendinous junction. Some lamellated corpuscles were found in the connective tissue of muscle septa and tendons, in contrast Ruffini corpuscles were not.

The receptive fields present in the rabbit rotator cuff. Filled circles are units with mechanical thresholds greater than 7 g, and open circles are units with mechanical thresholds less than 7 g.
Glenohumeral joint
Two studies using human cadavers have analysed the neurohistology of the shoulder joint. Guanche et al50 analysed three cadaveric human shoulders using gold chloride staining and light microscopy. They found that the superior, middle and inferior glenohumeral ligaments all contained mechanoreceptors (Golgi tendon organs, Ruffini's endings and Pacinian corpuscles) and free nerve endings. Only free nerve endings were identified in the biceps tendon and glenoid labral tissue. Hashimoto et al51 analysed 26 shoulder joints from 13 cadavers using immunohistochemistry and light microscopy. A small number of free nerve endings were identified in the innermost layer of the joint capsule. Many nerve fibres of large diameter were found in both the anteroinferior and posteriosuperior portions of the outermost layers of capsule (stratum fibrosum), while nerve fibres of various sizes were more numerous in posteriosuperior and anteroinferior portions of the boundary zone between the labrum and capsule in all layers.
Animal studies investigating the innervation of the shoulder joint have been relatively consistent in their findings. Backenkohler et al49 showed the Ruffini corpuscles were exclusively found in the ventral aspect of the joint capsule, while most lamellated corpuscles were located in the axillary portion of the joint capsule. Solomonow et al52 examined the distribution of neural elements in the glenohumeral capsule of cats. Again the greatest density of neural elements was found in the inferior aspect of the capsule; nerve fibres, free nerve endings, Golgi tendon organs, Ruffini's endings and Pacinian corpuscles were found in all areas of the capsule. Tarumoto et al53 studied the glenohumeral capsule in monkeys and again confirmed this dense inferior capsular innervation, particularly with Ruffini-like endings. They also found a high density of smaller peptidergic fibres that were presumed to be nociceptive, in the posterior half of the capsule; a small number of nociceptive fibres were found to innervate the marginal labrum. Overall, an increased density of mechanoreceptors in certain areas of the shoulder joint region coincides with zones where sensory control is most important because of increased biomechanical stress.54 The presence of corpuscular mechanoreceptors in the joint capsule and glenoid labrum shows their important role in inducing protective reflex actions in phases of extreme or abnormal movement.54 ,55
Subacromial bursa
The subacromial bursa (SAB) is an important structure that has been implicated in the pain of patients with rotator cuff tendinopathy and ‘impingement syndrome’. Human tissue from the SAB is readily available because of its frequent removal as part of ‘subacromial decompression’ (SAD) surgery. Numerous studies have shown that the SAB is richly innervated.56–58 Soifer et al57 examined cadaveric shoulder tissue using IHC and found that neural elements to be present in SAB, cuff tendon, biceps tendon and tendon sheath; the nerve supply to the SAB was significantly denser than that to the other tissues. Tomita et al58 examined tissue from the SAB of patients with varying degrees of cuff tendinopathy. They found that the SAB was innervated by large numbers of free nerve endings and mechanoreceptors (encapsulated corpuscles); they also found that there was a strong correlation between the density of neural elements at rest and shoulder pain at rest, those with higher densities of neural elements being more likely to experience pain at rest.
Ide et al56 investigated the innervation of the SAB in great detail using IHC and their results are shown in figure 5. A large number of C and Aδ fibres were found to innervate the SAB and they were shown to be immunoreactive to SP and CGRP, confirming their nociceptive role. A variety of mechanoreceptors were innervated by larger Aβ fibres and these fibres were not immunoreactive to SP. The Pacinian corpuscles were localised in the roof of the coracoacromial (C-A) arch side of the bursa, while the Ruffini endings were located at both the greater tuberosity and C-A side of the bursa; other unclassified mechanoreceptors were found in the bursa, some of which resembled Golgi-Mazzoni corpuscles. An intriguing study by Gotoh et al24 showed that patients with higher levels of SP in the SAB had greater levels of pain, and that SP levels were higher in non-perforated cuff tears versus perforated cuff tears.
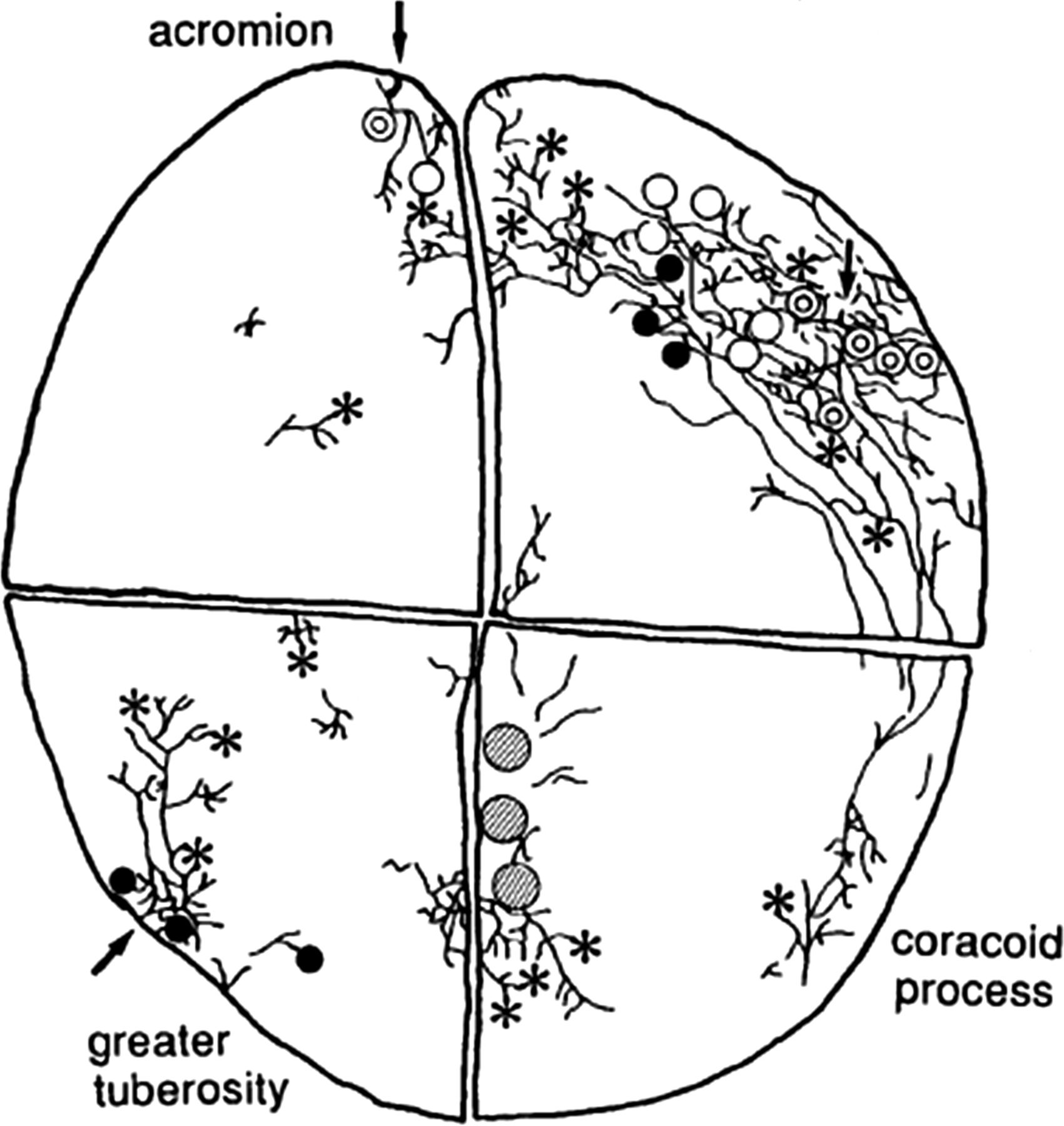
A schematic drawing of the major nerve fibre bundles and the distribution of sensory nerve endings in the SAB. Double circles indicate Pacinian corpuscles. Filled circles indicate Ruffini endings. Asterisks indicate free nerve ending plexus. Hatched areas indicate areas where cluster of Golgi-Mazzoni corpuscle-like receptors exists. White circles indicate type II unclassified receptors.
Long head of biceps tendon
The long head of biceps tendon (LHBT) is often implicated in shoulder pain and a degenerate LHBT frequently coexists with rotator cuff tendinopathy. Alpantaki et al59 analysed four cadaveric LHBT in humans using IHC. An extensive neural network was demonstrated along the tendon, with the densest innervation being present proximally and decreasing distally. This innervation was positive for SP and CGRP, suggesting the presence of thinly myelinated and unmyelinated sensory neurons; it was also positive for tyrosine hydroxylase, indicating the presence of post-ganglionic sympathetic fibres. Singaraju et al60 compared the neurohistology of the LHBT in patients undergoing surgery for pain related to degeneration with that of cadaveric specimens, none of which had evidence of cuff or LHBT degeneration. No difference was found in CGRP and SP staining between the two groups but there was a trend towards increased inflammation and vascularity in the surgical specimens. These results led the authors to conclude that neither the LHBT nor its sheath was the sole cause of anterior shoulder pain. Hashimoto et al51 showed that, in the central portion of the long head of biceps (LHB) tendon, relatively large nerve fibres inserted from the shoulder capsule. Tosounidis et al61 demonstrated by staining for the S-100 protein and neuropeptide Y that cells with neural differentiation within the LHBT were increased in acute injury and hypothesised that this provided a mechanism for LHBT-related pain. The function of the LHB tendon and its role in glenohumeral kinematics presently remain only partially understood62; likewise its role in shoulder pain remains controversial and incompletely understood.
C-A ligament
Konttinen et al63 demonstrated the C-A ligament to be aneural but the adjacent connective tissue and fat to be richly innervated; some of these nerves stained positively for Neuropeptide Y (sympathetic marker) and none for SP/CGRP. The rich innervation of the connective tissue adjacent to the coracoacromial ligament (CAL) is described above (figure 5). The aneural nature of the C-A ligament parenchyma has been subsequently confirmed by Tamai et al.64 Tamai et al64 also showed that the superficial periligamentous bursal tissue over the CAL was richly innervated with nociceptive fibres in patients with rotator cuff disease and not in controls; they hypothesised that this may be one of the causes of pain in this group of patients.
Neural anatomy of the shoulder
The nerves that contribute to the innervation of the anterior shoulder joint are the subscapular (C5/6), axillary (C5/6) and the lateral pectoral (C5/6) (figure 6).65–67 The subscapular nerves arise high from the posterior cord of the brachial plexus and supply a small portion of the anterior joint. The axillary nerve is the last nerve of the posterior cord of the brachial plexus before the latter becomes the radial nerve. As the axillary nerve courses across the subscapularis muscle, it releases its first articular branch as it runs across the anteroinferior capsule. The axillary nerve divides into two main branches at the lower aspect of subscapularis, the medial and lateral branches; the medial branch supplies the scapular side of the anteroinferior capsule, while the lateral branch supplies the humeral side of the anteroinferior capsule. The muscular branch which innervates teres minor, also issues a small articular branch at the level of the insertion of the long head of triceps. The lateral pectoral nerve arises from the lateral cord of the brachial plexus, passes superficial to the axillary vessels and pierces the clavipectoral fascia to reach the deep surface of pectoralis major. A small articular branch arises just before it pierces the fascia, this gives off branches to the coracoacromial (C-A) and coracohumeral (C-H) ligaments on its way laterally. Between these ligaments it then divides into two main branches; one passes beneath the C-A ligament to supply the SAB, the other passes across the C-A ligament to supply the anterior portion of the acromioclavicular joint.
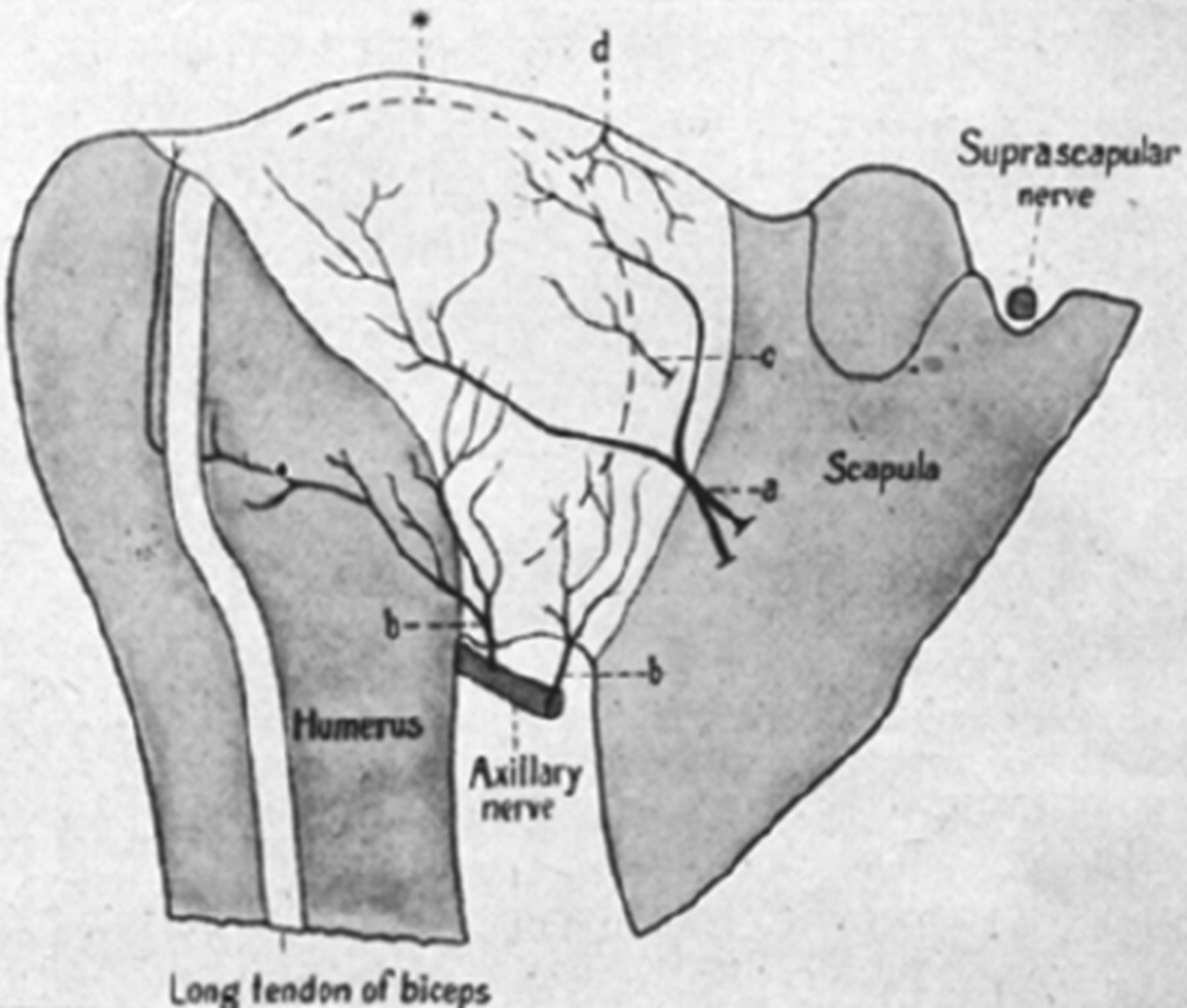
Profile reconstruction of the anterior side of right shoulder joint in a human fetus. *, Profile line of humeral head; a, articular nerve from the axillary and the musculocutaneous nerves; b, articular nerves from the axillary nerve; c, articular nerve from the subscapular nerve; d, branch of articular nerve from the suprascapular nerve.
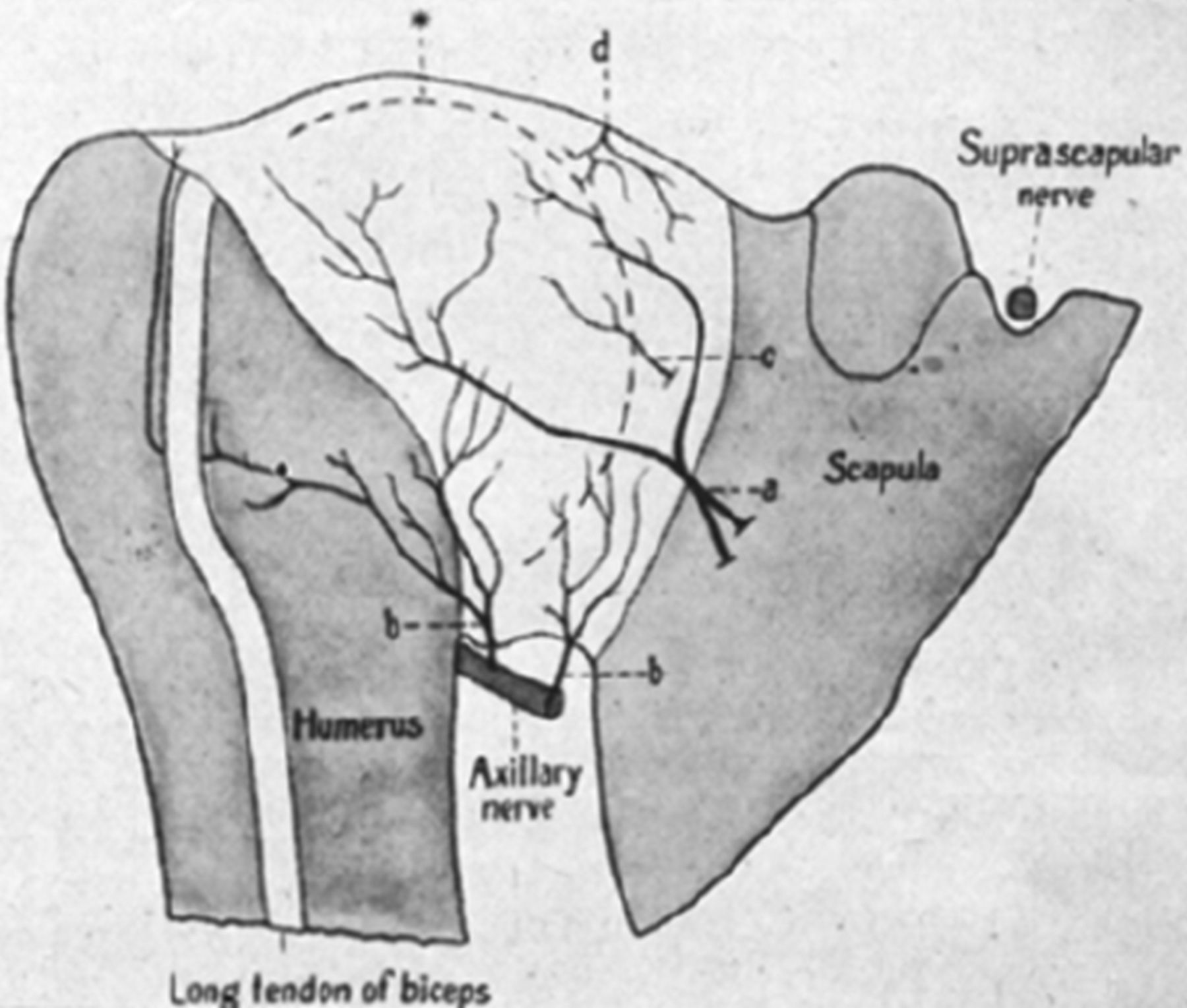
The nerves that contribute to the innervation at the posterior joint are the suprascapular nerve (SSCN) (C5/6) and axillary nerve (C5/6) (figure 7). The SSCN arises from the upper trunk of the brachial plexus. A large superior articular branch separates from the main nerve at an average of 4.5 cm proximal to the transverse scapular ligament,67 together these nerves pass underneath the transverse scapular ligament and enter the suprascapular notch. After entering the notch, the SSCN turns laterally and releases a small branch to the coracoclavicular ligaments. The main articular branch then advances laterally between the coracoid and supraspinatus, it divides into two terminal branches at this point; one descends to innervate the C-H ligament and the adjacent capsule, the other splits into several small branches which innervate the SAB and the posterior portion of the acromioclavicular joint capsule. The main SSCN passes into the suprascapular fossa where it releases a large muscular branch supplying the supraspinatus; at the level of the scapular spine a large inferior articular branch separates and travels obliquely to the posterior capsule. This inferior articular branch releases several branches which deviate upward and downward to terminate where the tendon of infraspinatus merges with the posterior capsule. The suprascapular nerve terminates by supplying infraspinatus. The contribution of the axillary nerve has been previously described. The sympathetic innervation of the shoulder joint is from the cervical ganglia (superior, middle and lower) via the grey rami communicantes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Profile reconstruction of posterior side of right shoulder joint in a human fetus. *, Profile line of humeral head; d, upper branch of the suprascapular nerve; e, lower branch of the suprascapular nerve; f, branches from nerves of supraspinatus and infraspinatus muscles; g, axillary branch to scapular part of capsule; h, axillary branches to humeral part of capsule.
Considering drivers of pain in the diagnosis and management of shoulder pain
The traditional triad of history, examination and investigations are used to guide the clinician towards a reasoned management plan for the patient's pain. It is also important to focus on features of the history and examination which hint at a greater degree of central sensitisation being present. A history of the pain radiating down the arm and the presence of ‘light touch’ hyperalgesia around the shoulder are both features of central sensitisation.38 Significant symptoms in the absence of radiological abnormality should ring alarm bells; in these cases treating an assumed peripheral pathology surgically may result in disaster as the majority of the pathology is in the higher pain processing systems.
The lack of reliable diagnostic tests for many shoulder pathologies68 including ‘impingement syndrome’69 ,70 reflects how subjective and variable clinical tests may be. This can be explained by mechanosensitivity changing as a result of any form of peri-articular inflammation with the majority of nociceptive fibres (C and δ) showing increased mechanosensitivity; indeed a large proportion of HTM are sensitised such that they respond to movements in the working range. This has been confirmed in the shoulder by Yamashita et al71 who showed that inflammation peripherally sensitised both low-threshold and high-threshold mechanoreceptors in a rabbit model. The classical inflammatory mediators such as bradykinin, the prostaglandins and 5-HT3 excite joint afferents and sensitise them to mechanical stimulation. Spinal cord neurons may be activated by noxious movements (NS, nociceptive specific) or they may respond with increasing intensity when the stimulus is increased from the innocuous to the noxious range (WDR, wide dynamic range). Both WDR and NS spinal cord neurons show enhanced responses to noxious stimuli applied in the inflamed joint, and NS neurons show a reduction in their mechanical threshold so that they now become excited by innocuous stimuli.
This means that any cause of inflammation in or around the shoulder may give rise to the so-called ‘irritable’ shoulder joint. Therefore, clinically it may be difficult to discriminate between rotator cuff tendinopathy and calcific tendinitis or inflammatory joint disease, for example; there is a clear lack of clarity with regard to whether common diagnostic tests used in clinical examination are useful in differentially diagnosing pathologies of the shoulder.68 ,72 Consequently, there has been the modern trend away from these bedside clinical tests towards radiological imaging modalities such as ultrasound and MRI.
Treatments of shoulder pain
The treatment of shoulder pain depends largely on the clinical diagnosis reached upon and it is beyond the scope of this article to deal with the specifics of all shoulder pathologies. We intend to focus on the aspects of treatment that are particularly relevant to pain processing in this overview. Before embarking upon a treatment, it is important to adequately assess the severity of the pain, as well as the impact of the pain upon the patient's shoulder function and quality of life; in this way any response to treatment can be more objectively measured and quantified. The visual analogue scale (VAS) is one of the many measures that can be used to assess pain,73 while there are numerous shoulder-specific scoring systems74 and quality-of-life measures.75 There are also more specific scoring systems which can help identify and quantify the neuropathic element of a patient's pain such as the pain DETECT questionnaire.76
Placebo effect
An important obstacle to measuring the effect of any treatment on pain is the significance of the placebo effect, which is ‘the patient response to inactive or sham treatments’. Placebo treatments have powerful analgesic effects and even placebo-controlled randomised trials may result in the efficacy of a treatment being misinterpreted.77 The DPMS is crucially important to placebo analgesia.78 A variety of factors including patient expectation, emotions such as anxiety and mood all have significant effects on the complex neurobiological process that lies behind placebo analgesia.79 ,80 Different types of neuropathic pain are affected in different ways by the placebo effect.81 While different treatments may have hugely differing strengths of placebo effect depending on multiple complex patient and treatment factors, it has been argued that the placebo effect of sham surgery is likely to be particularly strong.82 Consequently, the interpretation of treatment effects in shoulder pain is a highly contentious and controversial area. Maximising the placebo effect is useful for any clinician treating shoulder pain; ensuring that the patient has confidence in the treatment plan and allaying any anxieties are examples of ways in which this can be performed.
Pharmacotherapy
The World Health Organisation's analgesia ladder is frequently used to guide the treatment of pain and has been modified since its introduction in 1986.83 Simple agents such as Paracetamol and the non-steroidal anti-inflammatory drugs (NSAIDs) are generally first line. Paracetamol has both peripheral and central effects in inhibiting prostaglandin synthesis and activating the descending 5-HT system.84 The NSAIDs inhibit the cyclooxygenases (COX-1 and COX-2) to varying degrees resulting in both peripheral and central analgesic effects. Peripherally, they inhibit the prostaglandin-induced nociceptor sensitisation that occurs via TRPV1 and sodium channel activation; centrally, their effects are thought to be partly owing to the inhibition of the prostaglandin-mediated glycinergic neurotransmission.
Weak and strong opiates can then be added to the simpler agents in a step-wise fashion. The μ, δ, κ and opioid receptor-like-1 (ORL1) opioid receptors have been found in humans.4 The coupling of these receptors to potassium and sodium channels is believed to be the main mechanism by which endogenous and exogenous opioids produce analgesia. At a spinal level the most important mechanism of action is the presynaptic opioid receptor agonism which leads to a decrease in nociceptive afferent transmitter release; postsynaptic and interneuron opioid receptor agonism attenuates spinal cord outputs. Important sites of supraspinal opioid action are the PAG and RVM. Other drugs including certain antidepressants and anticonvulsants are particularly important in managing neuropathic pain.85 Table 2 summarises the classes of drugs that may be used in the management of acute and chronic shoulder pain. The decision-making process in analgesic pharmacotherapy involves the consideration of several complex factors including the nature and the chronicity of the pain; the evolving specifics relating to chronic neuropathic pain is a huge topic in its own merit.85 ,86
A summary of the analgesics that may be used in the management of shoulder pain
Injections and nerve blocks
A number of different injection sites have been used in treating shoulder pain including the glenohumeral joint, the subacromial space and several nerve blocks. Peripheral injections may be diagnostic, prognostic or therapeutic; they may consist of local anaesthetic with or without steroid. Local anaesthetics are sodium channel blockers and directly block the nerve conduction. The pain-relieving mechanisms of steroids are complex, not only do they have an anti-inflammatory effects, but they also appear to reduce nociceptor sensitivity and central sensitisation.87
The benefits of steroid injections have been demonstrated in both frozen shoulder and in cuff tendinopathy.88 ,89 However, the Cochrane review of the use of steroid injections for shoulder pain in frozen shoulder and in rotator cuff disease concluded that injections may be beneficial, but that their effect may be small and not well-maintained.90 This conflicting evidence reflects small trial sample sizes, variable methodological quality and a general heterogeneity.
Nerve blocks are widely used in managing intraoperative and postoperative pains in shoulder surgery; several nerve blocks or combinations of blocks are commonly used including the interscalene, suprascapular and axillary nerve blocks. The suprascapular nerve block has been shown to be more effective than subacromial infiltration or placebo after shoulder surgery.91 The suprascapular nerve block has also been shown to reduce pain in cases of frozen shoulder 1 month following an injection92 and in cases of general chronic shoulder pain of multiple aetiologies.93 Pulsed radiofrequency of the suprascapular nerve has also been reported as an effective treatment of chronic shoulder pain.94
Acupuncture
It is hypothesised that acupuncture relieves pain by activating Aδ and possibly C fibres via the mechanical stimulation of needles. Some trials have shown a benefit over placebo in treating shoulder pain,95 however the benefit could be attributed to the greater placebo effect of actual acupuncture over sham acupuncture. The Cochrane review regarding acupuncture for shoulder pain concluded that there was insufficient evidence to support or refute it as a treatment.96
Physiotherapy and activity modification
The proposed analgesic mechanisms of physiotherapy include rotator cuff muscle strengthening, augmenting scapulothoracic movement, increasing proprioceptive feedback and stretching tight structures. The exact nature of what shoulder physiotherapy consists of varies widely in the literature; for example, some regimes are patient-led, others more therapist-led, while some are particularly aggressive in terms of provoking pain symptoms. This has been emphasised by the Cochrane review which stated that the small sample sizes, variable methodological quality and heterogeneity in terms of population studied, physiotherapy intervention employed and length of the follow-up of randomised controlled trials of physiotherapy interventions results in little overall evidence to guide treatment.97 However, there is evidence to support the use of some interventions in specific cases,98 ,99 while physiotherapy has compared favourably with surgery in some trials.100 Stretching of the posteroinferior shoulder capsule is an effective treatment for shoulder pain in athletes with an associated internal rotation deficit.101 In this particular example the mechanisms behind the pain, the role of capsular receptors and the reason for pain improving with stretching remain unclear. Physiotherapy and activity modification frequently overlap, for example if pain is felt to be related to overuse then a reduction in activity levels may be followed by a supervised structured programme of rehabilitation. In some instances the individual may wish to accept that certain activities are no longer possible or that living with a certain level of pain is acceptable, this decision is often made having discussed all available treatment options with their physiotherapist, doctor or surgeon.
Surgery
Many of the surgical treatments of shoulder pain involve attempting to switch off the peripheral environmental triggers of pain by attempting to ‘fix’ the underlying ‘causal’ pathological process. Defining a causal pathological process is not always straightforward and this has been emphasised by the confusion surrounding the diagnosis and management of ‘impingement syndrome’.69 ,70 There remains a lack of high-quality evidence showing that surgery is effective in rotator cuff disease, for example102; the heterogeneous nature of patient groups and treatment methods make it hard to draw firm conclusions.103 The Cochrane review of surgery for glenohumeral osteoarthritis stated that there was a need for studies comparing shoulder surgery to sham, placebo and other non-surgical treatment options; the ethical and logistical difficulties conducting a trial that compared shoulder arthroplasty with sham surgery would be considerable. There is emerging evidence that certain surgical treatments such as rotator cuff repair result in significant sustained benefits in terms of pain and function.104 Certainly more large, high-quality randomised controlled trials are needed to answer these questions surrounding the effectiveness of surgery in specific shoulder pathologies.
The role of central sensitisation in shoulder pain has recently been explored in relation to ‘impingement syndrome’ and outcome after SAD.38 Patients with higher levels of punctate hyperalgesia and/or referred pain (features demonstrative of central sensitisation) did significantly worsen in terms of postoperative outcome. Central sensitisation has also been demonstrated in patients with hip and knee osteoarthritis.105 ,106 Gwilym et al105 demonstrated increased activity in the PAG with stimulation of the patient's skin in the areas of their referred arthritic pain. Joint arthroplasty can reverse pain-related changes in the thalamus in patients with hip and knee osteoarthritis, in association with their reduced pain and increased function following surgery.107 The normalisation of hyperaesthesia following knee joint arthroplasty has been demonstrated by Graven-Nielsen et al,37 implying that the central pain processes are maintained by peripheral input.
Discussion
The effective diagnosis and treatment of shoulder pain relies not only upon a detailed knowledge of the peripheral pathologies that may be present in the shoulder, but also a comprehensive understanding of how pain can be generated, propagated and modified in the human body. Human pain processing involves a far higher degree of neuronal plasticity than previously thought. Therefore, the location of receptors and the types of receptor present in the shoulder are of key importance in explaining this dynamic interaction between peripheral pathology and pain generation. However, there is still a long way to go in terms of fully understanding the variable relationship between peripheral pathology and perceived pain.
Our enhanced understanding of the molecular mechanisms involved in rotator cuff disease is resulting in the development of new treatment, for example the use of drugs and scaffolds to augment rotator cuff tendon repair.108 The role of the nervous system and specific neuropeptides in shoulder pain is still emerging, but undoubtedly furthering our understanding in this complex area has the potential to unlock some very powerful new diagnostic and therapeutic techniques. Genetic testing, biomarker measurement (both serum and peripheral) and pain-related bedside clinical tests are all examples of methods which can potentially be used to aid diagnosis and guide therapeutics. As the genetic component of shoulder pain is unravelled it may even be possible to determine which patients are particularly predisposed to problems so that they could be targeted with preventative treatments.
One new avenue for treatment is the pharmacological manipulation of the nervous system to modify the perception of pain. This may be performed peripherally, centrally or by a combination of the two. The early results of anti-NGF treatment of pain in knee osteoarthritis showed promise before trails were abandoned owing to serious neuropathic complications,109 demonstrating both the potential benefits and the pitfalls of new analgesic pharmacotherapies. SP has been implicated in the pathogenesis of tendinopathy in animal models110 ,111 and in humans.112 This raises the possibility of using SP antagonism to treat tendinopathy; in contrast, SP injections have shown an early promise in an animal model of tendon repair.113
The complex role of neuronal mediators like SP is demonstrated by its positive and negative effects in these studies, underlining how far we are from a full understanding of the dynamic role of the nervous system in painful musculoskeletal conditions. However, as this complexity is unravelled there is the potential for pharmacological interventions to be used in isolation or in combination with other treatments such as physiotherapy and surgery to create a multimodal approach which addresses all areas (peripheral and central) contributing to a patient's shoulder pain.
References
Footnotes
-
Contributors All authors have contributed to the drafting and finalisation of the manuscript.
-
Competing interests The authors of this work are funded by the Musculoskeletal Biomedical Research Unit of the National Institute for Health Research (BD, SG and AC), the Jean Shanks Foundation (BD) and the Lord Nuffield Scholarship for Orthopaedic Surgery (BD).
-
Provenance and peer review Not commissioned; externally peer reviewed.