

Abstract
Objective. To assess the effect of B cell depletion therapy on effector CD4+ T cell homeostasis and its relation to objective measures of disease activity in patients with primary Sjögren syndrome (pSS).
Methods. Twenty-four patients with pSS treated with rituximab (RTX) and 24 healthy controls (HC) were included. Frequencies of circulating effector CD4+ T cell subsets were examined by flow cytometry at baseline and 16, 24, 36, and 48 weeks after the first RTX infusion. Th1, Th2, follicular Th (TFH), and Th17 cells were discerned based on surface marker expression patterns. Additionally, intracellular cytokine staining was performed for interferon-γ, interleukin (IL)-4, IL-21, and IL-17 and serum levels of these cytokines were analyzed.
Results. In patients with pSS, frequencies of circulating TFH cells and Th17 cells were increased at baseline compared with HC, whereas frequencies of Th1 and Th2 cells were unchanged. B cell depletion therapy resulted in a pronounced decrease in circulating TFH cells, whereas Th17 cells were only slightly lowered. Frequencies of IL-21–producing and IL-17–producing CD4+ T cells and serum levels of IL-21 and IL-17 were also reduced. Importantly, the decrease in circulating TFH cells was associated with lower systemic disease activity over time, as measured by the European League Against Rheumatism Sjögren’s Syndrome Disease Activity Index scores and serum IgG levels.
Conclusion. B cell depletion therapy in patients with pSS results in normalization of the elevated levels of circulating TFH cells. This reduction is associated with improved objective clinical disease activity measures. Our observations illustrate the pivotal role of the crosstalk between B cells and TFH cells in the pathogenesis of pSS.
To date, the immunopathogenetic mechanism of primary Sjögren syndrome (pSS) has not been fully elucidated. Both T cells (predominantly CD4+ T cells) and B cells infiltrate the salivary glands. This inflammatory response is accompanied by loss of glandular architecture and impaired function of the gland1. B cell hyperactivity is a prominent feature of the disease, which is manifested by hyperimmunoglobulinemia, the presence of anti-SSA/Ro and anti-SSB/La autoantibodies, and the significantly increased risk for development of mucosa-associated lymphoid tissue B cell lymphoma2, among other manifestations. B cells are therefore thought to play a critical role in immunopathogenesis of pSS.
In addition to B cells, different CD4+ T cell subsets may be involved in pSS pathogenesis. Th1 cells and Th17 cells seem to infiltrate the glands at an early disease stage, as evidenced by detection of interferon (IFN)-γ and interleukin (IL)-17, whereas infiltration of follicular Th (TFH) cells is associated with later stages3. TFH cells are involved in formation of germinal centers (GC) and generation of (autoreactive) plasma cells4. Because the frequency of circulating TFH cells is increased in patients with pSS and given their role in B cell activation, TFH cells may play a prominent role in the establishment and maintenance of B cell hyperactivity in pSS5,6.
B cell hyperactivity may be targeted by B cell depletion therapy with rituximab (RTX), an anti-CD20 monoclonal antibody. RTX seems a beneficial treatment modality for pSS patients with moderate to high systemic disease activity7,8,9,10, although this was not confirmed in a large randomized trial, nor in patients with longstanding disease11,12. It is therefore important to understand how and in which subgroup of patients RTX treatment may improve clinical outcome. B cell depletion therapy with RTX results in only a moderate drop in serum IgG levels8,11,13, while long-lived IgA- or IgG-producing cells persist in the parotid glands14. Thus, also B cell functions other than antibody production are probably implicated in the mode of action of B cell depletion therapy in pSS. These functions may comprise antigen presentation and secretion of various regulatory and proinflammatory cytokines15,16. Cytokine secretion by B cells may affect the differentiation of naive CD4+ T cells into effector CD4+ T cell subsets. In turn, CD4+ T cells may subsequently influence the activity of B cells17,18. We hypothesize that this crosstalk plays a critical role in the pathogenesis of pSS. To test this hypothesis, we assessed the effect of B cell depletion therapy with RTX on the distribution and in vitro cytokine-producing capacity of circulating effector CD4+ T cell subsets in patients with pSS and its relation to objective measures of disease activity.
MATERIALS AND METHODS
Study population and procedures
For our present study, patient material was used from our previously reported open-label study on RTX, which was an extension of our placebo-controlled study8. The extension study included 28 patients with pSS13. Twenty-four patients were analyzed in the current study; 4 patients were not included because of the unavailability of stored peripheral blood mononuclear cell (PBMC) samples (n = 3) or serum sickness-like manifestations after the first dose of RTX (n = 1). All patients fulfilled the revised American-European Consensus Group criteria for pSS19. Eighteen of these 24 patients were treated before with RTX during previous studies and received their second (n = 13), third (n = 3), or fourth (n = 2) course of treatment; the remaining 6 patients were RTX-naive. For the patients who were re-treated with RTX, re-treatment started at least 1 year after the last course, i.e., after reappearance of circulating B cells to baseline levels and after recurrence of symptoms. Parotid gland biopsies were available from all patients before treatment and in addition from 3 patients at 16 weeks after treatment. Patient characteristics at baseline are presented in Table 1. Patients were treated on days 1 and 15 with intravenous (IV) injections of 1000 mg RTX (Roche). To minimize the risk of infusion reactions and serum sickness, all patients were treated with methylprednisolone (100 mg IV), acetaminophen (1000 mg orally), and clemastine (2 mg IV) prior to each infusion and received oral prednisone for 5 days after each infusion. Except for these precautionary medications, treatment with concomitant immunosuppressants was discontinued, including hydroxychloroquine and oral prednisone.
Baseline characteristics of patients with primary Sjögren syndrome treated with rituximab. Results are expressed as mean ± SD or median (IQR), unless otherwise indicated.
PBMC were isolated at baseline and 16, 24, 36, and 48 weeks after the first dose of RTX in the current study design. At the same timepoints, objective signs of disease activity were measured, including the European League Against Rheumatism Sjögren Syndrome Disease Activity Index (ESSDAI)20, ESSDAI without the biological domain (clinESSDAI)21, serum IgG levels, and stimulated whole saliva production. Twenty-four age- and sex-matched healthy individuals were included in the study as controls (HC). All patients and HC provided informed consent and the study was approved by the local Medical Ethics Committee of the University Medical Center Groningen (METc2008.179).
Analysis of effector CD4+ T cell subsets by flow cytometry: surface marker analysis
Effector CD4+ T cell subsets can be further divided into 4 subsets by using a combination of different chemokine receptors22,23. Accordingly, we defined Th1, Th2, Th17, and TFH cells as CD45RA− FoxP3− CXCR5− CXCR3+ CCR4− CCR6–, CD45RA− FoxP3− CXCR5− CXCR3− CCR4+ CCR6–, CD45RA− FoxP3− CXCR5− CXCR3− CCR4+ CCR6+, and CD45RA− FoxP3− CXCR5+ PD-1+ CD4+ T cells, respectively (described in Supplementary Figure 1, available with the online version of this article). For flow cytometry analysis, cryopreserved PBMC from 24 patients with pSS and 24 HC were stained for surface markers with antibodies listed in Supplementary Table 1, available with the online version of this article. Subsequently, cells were stained with a fixable viability dye (eBioscience), fixed and permeabilized in Fix/Perm buffer and stained with anti-FoxP3 (eBioscience). Fluorescence-minus-one control stains were included to determine background levels of staining. After staining, cells were washed and immediately analyzed by flow cytometry.
In vitro cytokine production analysis
In addition to surface marker analysis, in vitro stimulation and intracellular cytokine staining was performed using PBMC from 20 consecutive patients with pSS and 11 consecutive HC. Cryopreserved PBMC were thawed and stimulated in vitro with 5 ng/ml phorbol myristate acetate (Sigma-Aldrich) and 0.17 µg/ml calcium ionophore A23187 (Ca-I; Sigma-Aldrich) for 16 h at 37°C and 5% CO2. These conditions were tested as optimal for analysis of in vitro cytokine production by T cells after thawing. As a negative control, 1 sample of each cell suspension remained without stimulation. Directly after addition of the stimulants, 10 µg/ml brefeldin A (Sigma-Aldrich) was added. After stimulation, cells were fixed and permeabilized in Fix/Perm buffer (eBioscience) for 45 min at 4°C. Then, samples were labeled by fluorescent cell barcoding (FCB)24. After washing and permeabilization, different FCB-labeled samples were combined into 1 FACS tube and stained for 1 h with antibodies listed in Supplementary Table 1, available with the online version of this article. After staining, cells were washed and analyzed by flow cytometry for intracellular production of IFN-γ, IL-4, IL-17, and IL-21, which are 4 signature cytokines of Th1, Th2, Th17, and TFH cells, respectively. Because stimulation reduces surface expression of CD4 on T cells, cytokine-producing CD4+ T cells were identified indirectly by gating on CD3+CD8− lymphocytes. Unstimulated samples were used to delineate positive and negative populations and results are expressed as the percentage of single cytokine-producing cells within the total CD4+ T cell population. Dead cells not washed out by several washing steps were excluded on the basis of their forward- and side-scatter profile.
For all flow cytometry analyses, data were acquired using a FACS-LSRII flow cytometer (Becton Dickinson) and analyzed using the Kaluza 1.2 Flow Analysis software (Beckman Coulter) or FlowJo analysis software (Tree Star).
Absolute cell count in blood samples
In patients with pSS, absolute numbers of circulating CD4+ T cells and CD19+ B cells were measured in fresh blood samples by using the MultiTest TruCount method (Becton Dickinson), as described by the manufacturer. These numbers were used to convert the frequencies of different CD4+ T cell subsets into absolute numbers.
Measurement of serum cytokines and autoantibodies
Serum levels of IL-21, IL-17, IFN-γ, and IL-4 were measured using a ProcartaPlex human cytokine immunoassay (eBioscience), according to the manufacturer’s protocol. Serum levels of anti-SSA/Ro and anti–SSB/La at baseline and Week 16 were measured with a Phadia 250 analyzer using EliA Ro and EliA La (Phadia). Levels were expressed in arbitrary units per milliliter (AU/ml).
Quantitative PCR and immunofluorescence for detection of glandular IL-21 expression
Total RNA was extracted from microdissected infiltrated regions of frozen parotid gland tissue, using the RNeasy Micro Kit (Qiagen). cDNA was subsequently synthesized. Primers for β-actin (forward 5′-GAG CGG GAA ATC GTG CGT GAC-3′ and backward 5′-AGG AAG GAA GGC TGG AAG AGT GC-3′) and IL-21 (forward 5′-AAG CTC CCA AGG TCA AGA TCG-3′ and backward 5′-AGC AGG AAA AAG CTG ACC ACT-3′) were used for SYBR Green-based real-time quantitative PCR analysis (Bio-Rad). The mean of triplicate results was normalized to expression of endogenous transcripts encoding for β-actin to calculate relative mRNA expression of IL-21. Fold change values were determined using the 2^(ΔΔCt) method. The median baseline Ct value was used as calibrator. For analysis of IL-21 protein expression, formalin-fixed, paraffin-embedded sections (4 μm) of parotid gland tissue were incubated with primary antibody to IL-21 overnight at 4°C (NBP1-02706, Novus Biologicals). Prior to incubation, heat-induced epitope retrieval was performed using citrate buffer (pH 6). Anti–IL-21 antibodies were visualized using anti-rabbit-Af594-conjugated antibody (Invitrogen).
Statistical analysis
Statistical analysis was performed using SPSS Statistics 20 software (IBM). Results are presented as mean ± SD or median (interquartile range), for normally and non-normally distributed data, respectively. The Mann-Whitney U test was used to compare differences in frequencies of effector CD4+ T cell subsets between patients and HC. Generalized estimating equations (GEE) with exchangeable correlation structure were used to analyze the presence of different T cell subsets over time within subjects. If residuals were non-normally distributed, variables were square-root or log transformed before entering into the equation. We have previously reported that B cells started to reappear at Week 24 after RTX treatment25. Therefore, data from baseline up to Week 24 were used to assess change over time compared with baseline, i.e., during B cell depletion. Data from Week 24 up to Week 48 were used to assess change over time compared with Week 24, i.e., during B cell repopulation. Associations between the development of different objective disease variables and T cell subsets were also tested with GEE. Two-tailed p values < 0.05 were considered statistically significant.
RESULTS
Frequencies of circulating TFH cells and Th17 cells, defined by surface marker expression, are increased in patients with pSS compared with HC
To study homeostasis of CD4+ T cell subsets with acquired effector function in patients with pSS, we analyzed the relative distribution of CD4+ CD45RA− T cell subsets, i.e., antigen-experienced memory CD4+ T cells. Analysis of surface marker expression revealed that frequencies of CD4+ CD45RA− FoxP3− CXCR5+ PD-1+ cells were increased in patients with pSS compared with HC (Figure 1A; p < 0.001). These CD4+ CD45RA− CXCR5+ PD-1+ cells can be considered as circulating counterparts of TFH cells in lymphoid tissue, because they share phenotypical and functional characteristics26,27. We therefore refer to them as circulating TFH (cTFH) cells. Frequencies of Th17 cells were also increased in patients with pSS (Figure 1B; p = 0.009). Of note, there was a positive correlation between frequencies of cTFH and Th17 cells (Spearman’s ρ = 0.44; p = 0.031). No significant differences were observed in frequencies of Th1 cells or Th2 cells between patients with pSS and HC (Supplementary Figure 2, available with the online version of this article).

Frequencies of TFH cells and Th17 cells are increased at baseline in patients with pSS. Frequencies of circulating (A) TFH cells (CD45RA− CXCR5+ PD-1+) and (B) Th17 cells (CD45RA− CXCR5− CXCR3− CCR4+ CCR6+) in patients with pSS (n = 24) and HC (n = 24) are displayed. Additionally, frequencies of circulating (C) IL-21 and (D) IL-17−producing CD4+ T cells in patients with pSS (n = 20) and HC (n = 11) are displayed. Horizontal lines indicate the median. P value < 0.05 was considered statistically significant. P values were calculated using the nonparametric Mann-Whitney U test. TFH: follicular Th; pSS: primary Sjögren syndrome; HC: healthy controls; IL: interleukin.
Analysis of effector subsets by surface marker expression does not necessarily reflect cytokine-producing capabilities of these cells. We therefore subsequently analyzed the production of IL-21, IL-17, IFN-γ, and IL-4 by in vitro stimulated CD4+ T cells from patients with pSS and HC. Despite the observation that frequencies of surface marker–defined cTFH cells and Th17 cells were elevated, patients with pSS did not display aberrant frequencies of IL-21–producing or IL-17–producing CD4+ T cells at baseline, compared with HC (Figure 1C–D). Also no substantial alterations in frequencies of CD4+ T cells producing IFN-γ or IL-4 were observed in patients with pSS, compared with HC (Supplementary Figure 2, available with the online version of this article). For all surface marker–defined and cytokine-defined CD4+ T cell subsets, no significant differences were observed between RTX-naive patients and patients who had received RTX previously (data not shown).
Frequencies of circulating TFH cells (and to a small extent also Th17 cells) are reduced by B cell depletion therapy
Next, the effect of B cell depletion therapy with RTX on circulating effector CD4+ T cell subsets was evaluated. B cell depletion therapy resulted in a decrease of almost 50% in frequencies of cTFH cells (Figure 2A; p < 0.001). At Week 24, frequencies of cTFH cells in patients with pSS reached levels comparable to HC. Although Th17 cell frequencies did not decrease at the population level, a small but significant decrease in individual patients over time was observed by GEE analysis (Figure 2B; p = 0.041). No changes in frequencies of Th1 and Th2 cells were observed (Supplementary Figure 3, available with the online version of this article). During B cell repopulation, frequencies of cTFH cells returned to baseline levels of the patients (Figure 2A; p = 0.042). Analysis of absolute numbers of effector CD4+ T cell subsets during B cell depletion revealed that cTFH cells decreased significantly, whereas Th17, Th1, and Th2 cells were not significantly altered (Supplementary Table 2).

Predominantly TFH cells decrease after rituximab treatment. Frequencies are displayed of circulating (A) TFH cells (CD45RA− CXCR5+ PD-1+) and (B) Th17 cells (CD45RA− CXCR5− CXCR3− CCR4+ CCR6+) during B cell depletion (week 0–24) and B cell repopulation (week 24–48). Horizontal lines (black) indicate the median. Median blood CD19+ B cell numbers are shown in grey. The development of frequencies within subjects over time during B cell depletion was analyzed using generalized estimating equations. Frequencies within subjects during B cell repopulation were analyzed using the Wilcoxon matched pairs test. P value < 0.05 was considered statistically significant. TFH: follicular Th.
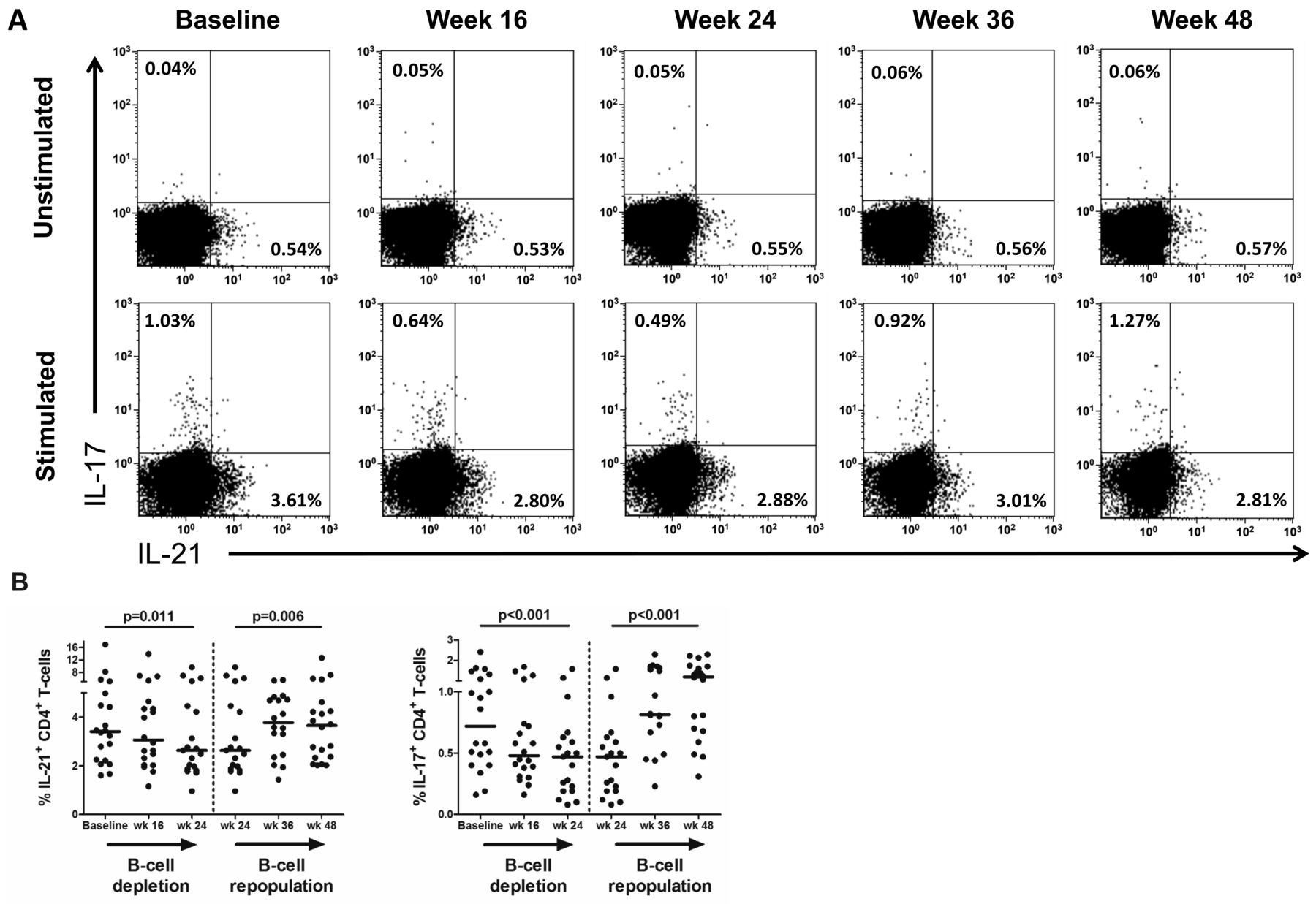
Frequencies of IL-21–producing and IL-17–producing CD4+ T cells are also reduced by B cell depletion therapy
Although frequencies of both IL-21–producing and IL-17–producing CD4+ T cells were not significantly altered in patients with pSS at baseline, compared with HC, these cytokine-producing cells were significantly affected by B cell depletion therapy (Figure 3A). Both frequencies of IL-21−producing CD4+ T cells (Figure 3B; p = 0.011) and IL-17−producing CD4+ T cells (Figure 3B; p < 0.001) were reduced during B cell depletion. In contrast, no substantial alterations in frequencies of circulating CD4+ T cells producing IFN-γ or IL-4 were observed (Supplementary Figure 3, available with the online version of this article). During B cell repopulation, frequencies of IL-21–producing and IL-17−producing CD4+ T cells increased again to levels present at baseline (Figure 3B). Analysis of absolute numbers of CD4+ T cells producing IL-17 during B cell depletion and repopulation gave comparable results. Absolute numbers of IL-21− producing CD4+ T cells were not significantly affected by B cell depletion, although a trend toward lower numbers was present (Supplementary Table 2).
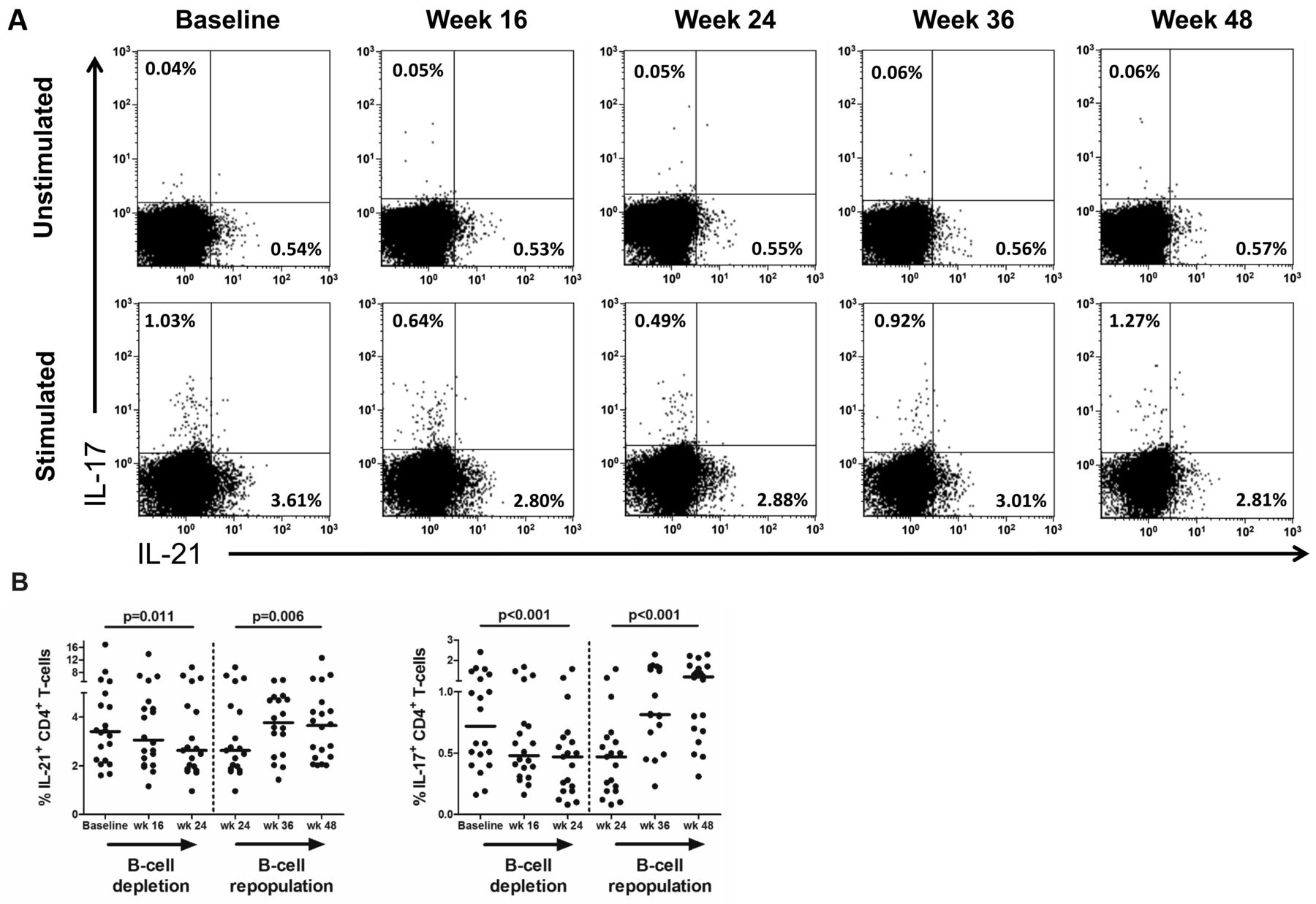
In vitro IL-21 and IL-17 production by CD4+ T cells after rituximab (RTX) treatment. A. Representative flow cytometry plots of in vitro IL-21 and IL-17 production in circulating CD4+ T cells from 1 patient with pSS at baseline and at 16, 24, 36, and 48 weeks after the first dose of RTX. The plotted cells were gated as CD3+CD8− T cells and gate settings were based on the unstimulated sample (biological control). Values in each gate represent the percentages of positive cells. B. In vitro cytokine production by CD4+ T cells in 20 patients with pSS. Frequencies of IL-21–producing and IL-17–producing CD4+ T cells are displayed. Frequencies within subjects over time were analyzed with generalized estimating equations during B cell depletion (week 0–24) and B cell repopulation (week 24–48). Horizontal lines indicate the median. P value < 0.05 was considered statistically significant. IL: interleukin; pSS: primary Sjögren syndrome.
Serum IL-21 and anti–SSA/-SSB levels are lowered by B cell depletion therapy
Because we observed that cTFH cells were reduced by B cell depletion therapy, and given the observation that treatment results in lowering of total IgG levels (± 15%)13, we analyzed whether serum levels of IL-21, anti-SSA/Ro, and anti-SSB/La were also affected. Serum IL-21 was significantly lowered by B cell depletion (Supplementary Figure 4, available with the online version of this article; p < 0.001). Importantly, decrease in serum IL-21 was associated with the reduction in cTFH cells over time (p < 0.001). Serum levels of IL-17, IFN-γ, and IL-4 were detectable at low levels in 58%, 92%, and 42% of the patients at baseline, respectively. During B cell depletion, in patients with detectable levels at baseline, a decrease in IL-17 and IL-4 was observed, whereas IFN-γ levels remained unchanged (Supplementary Figure 4). Anti-SSA/Ro titers were decreased by about 25% at Week 16, compared with baseline [8498 (5989–13009) AU/ml at Week 16; 11312 (7196–18254) AU/ml at baseline; n = 24; p < 0.001]. A similar reduction of ± 25% was observed for anti-SSB/La titers [249 (80–4966) AU/ml at Week 16; 339 (140–6886) AU/ml at baseline; n = 18; p < 0.001]. In addition to serum levels of IL-21, mRNA and protein expression levels of IL-21 were analyzed in parotid gland tissue of 3 patients at baseline and 16 weeks after treatment. Both mRNA and protein expression tended to decrease after treatment (Supplementary Figure 5, available with the online version of this article). These data suggest that not only numbers of cTFH cells are affected by B cell depletion therapy, but also their functionality in terms of IL-21 production, likely also locally in the inflamed salivary glands, resulting in less B cell help.
Reduced frequencies of circulating TFH cells and Th17 cells are associated with improvement in objective measures of disease activity and autoantibody titers
Because cTFH and Th17 cells, as well as IL-21–producing and IL-17–producing CD4+ T cells were affected by B cell depletion therapy, we subsequently analyzed associations between changes in these subsets and objective measures of disease activity over time (Table 2). Importantly, the decrease in frequencies of cTFH cells was significantly associated with amelioration of systemic disease activity, as measured by ESSDAI (p < 0.001) and clinESSDAI (p = 0.001), and with a decrease in total IgG (p < 0.001), anti-SSA/Ro (p < 0.001), and anti-SSB/La (p < 0.001) in serum. Reduced frequencies of IL-21–producing CD4+ T cells were associated only with improvement in ESSDAI (p < 0.001). The course of cTFH cells or IL-21–producing CD4+ T cells during B cell depletion did not significantly differ between patients with (n = 7) and without (n = 15) GC in parotid gland tissue at baseline. Frequencies of IL-17–producing CD4+ T cells were significantly associated with levels of IgG (p = 0.001) and anti-SSB/La (p < 0.001) over time. Further, we found that lower Th17 cell frequencies were associated with higher stimulated whole saliva production within patients over time (p < 0.001). cTFH cells tended to be associated with stimulated saliva production as well (p = 0.007).
Associations between clinical or laboratory variables and effector CD4+ T cell subsets within patients during B cell depletion.
DISCUSSION
Here, we show by surface marker analysis that frequencies of cTFH cells (CD4+ CD45RA− FoxP3− CXCR5+ PD-1+) and Th17 cells (CD4+ CD45RA− FoxP3− CXCR5− CXCR3− CCR4+ CCR6+) are increased in patients with pSS compared with HC, while Th1 and Th2 cell frequencies are unchanged. We further show that B cell depletion therapy reduces the elevated frequencies of cTFH cells and to a smaller extent also Th17 cells. In addition to the effects on surface marker–defined cTFH and Th17 cells, a specific reduction of IL-21–producing and IL-17–producing CD4+ T cells is seen after B cell depletion therapy. This is further reflected by lower serum levels of IL-21 and IL-17 after treatment. When circulating B cells repopulate, the observed effects on these pathogenic T cell subsets are reversed. Importantly, reductions in cTFH cells and to a lesser extent also Th17 cells are associated with amelioration of several objective disease activity variables.
Our findings confirm previous observations that levels of cTFH cells and Th17 cells, as defined by surface marker expression, are increased in patients with pSS, compared with HC5,6,26,28. We also observed that frequencies of cTFH cells and Th17 cells at baseline are correlated. This might be because differentiation pathways for TFH cells and Th17 cells are interconnected, mediated by IL-6 and IL-214,29,30. Although we observed an increase in both cTFH cells and Th17 cells in patients with pSS at baseline, this is not reflected by a similar increase in IL-21–producing or IL-17−producing CD4+ T cells. For IL-17–producing CD4+ T cells, comparable levels between patients with pSS and HC were previously noted31,32. Apparently, at least for cTFH cells and Th17 cells, there is no clear association between surface marker expression and signature cytokine production by these subsets in patients with pSS. The lack of correlation may be explained in 2 different ways that are not mutually exclusive. First, it is known that cytokine production is not limited to 1 subset, because not only CD45RA− CD4+ CXCR5+ PD-1+ cells, but also CD4+ CXCR5+ PD-1− cells and CD4+ CCR9+ T cells are able to produce IL-2123,33. Accordingly, IL-17 can be produced by all CD4+ CCR6+ cells, including Th17 cells (CD45RA− CD4+ CCR4+ CCR6+ T cells)34. Second, not all effector subsets in the circulation are yet fully capable of cytokine production, and additional signals (e.g., proinflammatory cytokines) in the inflamed tissue are necessary for these cells to become cytokine-producing effector CD4+ T cells. Dependency on local cytokine signals that drive cytokine production by effector CD4+ T cells has been described for IL-17−producing CD4+ T cells34. Likely, in patients with pSS, both cTFH and Th17 cells start to produce cytokines only upon arrival in glandular tissue. Accordingly, in minor salivary glands of patients with pSS, IL-17–producing CD4+ T cells are enriched in the lymphocytic infiltrate, compared with sicca controls35. Further, IL-21 and IL-17 are significantly upregulated in minor salivary gland tissue of patients with pSS, both at protein and mRNA level35,36,37,38. Local production of cytokines may also underlie the reported increase in serum levels of IL-21 and IL-1737,39,40,41,42. We therefore suggest that assessment of circulating effector subsets by surface marker expression provides better prognostic value, compared with measurement of in vitro cytokine production by circulating CD4+ T cells.
Importantly, we demonstrated in this study that B cell depletion therapy reduces both the levels of cTFH cells and IL-21–producing CD4+ T cells in patients with pSS. Both effects are reversed during B cell repopulation. This observation indicates that the drop in cTFH cells is a specific effect of RTX and is not, for example, due to administration of methylprednisolone, used to prevent infusion reactions. Previous studies in type 1 diabetes and granulomatosis with polyangiitis have also shown that B cell depletion therapy affects cTFH cells42,43. However, the immunopathology of these diseases is significantly different from pSS and these studies have not related their findings to changes in disease activity. In the current study, we observed that reduced cTFH cell frequencies in patients with pSS after B cell depletion are strongly associated with amelioration of several objective measures of disease activity, including ESSDAI, clinical ESSDAI, serum IgG, anti-SSA/Ro, anti-SSB/La, and to a lesser extent also stimulated saliva production.
Because TFH cells are important for GC formation and plasma cell differentiation4, reduced numbers of TFH cells may lead to less T cell–dependent B cell activation of persisting B cells in the salivary glands. We assume that the observed reduction in cTFH cells reflects the overall reduction of TFH cell activity in the body, because others have shown that the proportion of cTFH cells correlates well with TFH cell activity, including autoantibody production26,27. Preliminary data in 3 patients indicate that glandular TFH cell activity in patients with pSS, as reflected by the presence of IL-21–producing cells and total levels of IL-21 mRNA, is indeed decreased by B cell depletion therapy, whereas the total number of infiltrating T cells remained unaffected by treatment44. Reduction of TFH cells in patients with pSS may contribute to the decline in GC activity that is observed in parotid glands of patients with pSS after B cell depletion, even though the glands still contain appreciable numbers of B cells44. The combination of reduced TFH cells and depletion of CD20+ plasma cell precursors likely results in decreased generation of plasmablasts and short-lived plasma cells, which may explain lowering of anti-SSA/Ro60 antibodies that are partly produced by short-lived plasma cells45. However, probably not all plasmablasts are affected by treatment, because Dass, et al have shown using high-sensitivity flow cytometry that low numbers of circulating plasmablasts can still be present after B cell depletion therapy46. Although we did not analyze circulating plasmablasts in the current study, the decrease in serum IgG levels and autoantibodies after treatment suggests that at least a significant fraction of (autoreactive) plasma cell precursors is affected by treatment. B cell depletion therapy thus acts as a 2-edged sword: it results in fewer B cells that can be activated in the glands and in fewer TFH cells that can activate the persisting B cells. Also, circulating IL-17−producing CD4+ T cells are reduced after treatment, and to a minimal extent also circulating Th17 cells. The reduction in circulating IL-17–producing CD4+ T cells is reflected in the target tissue, because the amount of IL-17 protein in minor salivary gland tissue of patients with pSS is reduced after B cell depletion therapy35. Our observation that levels of circulating Th17 cells correlate with stimulated saliva production over time provides additional evidence for the pathogenic potential of Th17 cells in the glands.
The reduction in cTFH cells and Th17 cells might be explained by direct and/or indirect effects of B cell depletion. A minor fraction of IL-17–producing CD4+ T cells in peripheral blood from patients with pSS expresses low levels of CD2047. Binding of RTX to CD20 may cause a direct loss of these cells. In addition, indirect effects caused by the depletion of B cells may contribute to the decrease in Th17 cells, and in particular, TFH cells. Development of TFH cells, and to a lesser extent also Th17 cells, is dependent on the presence of IL-64,30. B cells, and more specifically, plasmablasts are potent producers of this cytokine48. In line with this notion we have shown that B cell depletion therapy results in decreased serum concentrations of IL-6 in patients with pSS39. Depletion of B cells by RTX may therefore reduce IL-6 mediated development of TFH cells and Th17 cells. IL-6 also stimulates the secretion of IL-21 and IL-17 by these T cell subsets4, which may explain the reduced production of IL-21–producing and IL-17–producing CD4+ T cells, and lowering of IL-21 and IL-17 levels in serum. The dependence of IL-17 production on (IL-6–producing) B cells is further illustrated by studies in mice and humans, which show that specifically Th17 activation, reflected by IL-17–production, is impaired by depletion of (IL-6–producing) B cells49,50. Th1 and Th2 cells are not dependent on IL-6 for their differentiation, and this may explain why these subsets are not affected by B cell depletion therapy.
Further studies are necessary to provide direct evidence that the ablation of IL-6 producing B cells is responsible for the observed effects of B cell depletion on CD4+ T cell subsets. The strong association between cTFH cells and clinESSDAI illustrates the central role of these cells in pathogenesis of pSS. We therefore hypothesize that in particular, patients with active systemic disease are likely to benefit from B cell depletion therapy. We further suggest that the crosstalk between B cells and CD4+ T cells is an appropriate target for therapy in pSS.
ONLINE SUPPLEMENT
Supplementary material accompanies the online version of this article.
Acknowledgment
The authors thank Rudi Hendriks for sharing his expertise on surface marker expression experiments and Marjolein de Bruijn for excellent technical assistance.
Footnotes
This investigator-driven study was financially supported by Roche, Woerden, the Netherlands, which supplied the study medication. There was no involvement of the funding source in study design or patient recruitment, or data collection, analysis, and interpretation, or writing of the report.
- Accepted for publication October 18, 2016.








{kind=link}
{kind=link}
{kind=link}