Article Text

Abstract
In recent years, the activation of the insulin-like growth factor (IGF) system in cancer has emerged as a key factor for tumour progression and resistance to apoptosis. Therefore, a variety of strategies have been developed to block the type I IGF receptor (IGF-I-R), which is thought to mediate the biological effects of both IGF-I and IGF-II. However, recent data suggest that the IGF signalling system is complex and that other receptors are involved. To unravel the complexity of the IGF system in thyroid cancer, IGF-I and IGF-II production, and the expression and function of their cognate receptors were studied. Both IGFs were found to be locally produced in thyroid cancer: IGF-I by stromal cells and IGF-II by malignant thyrocytes. Values were significantly higher in malignant tissue than in normal tissue. IGF-I-Rs were overexpressed in differentiated papillary carcinomas but not in poorly differentiated or undifferentiated tumours, whereas insulin receptors (IRs) were greatly overexpressed in all tumour hystotypes, with a trend for higher values in dedifferentiated tumours. As a consequence of IR overexpression, high amounts of IR/IGF-I-R hybrids (which bind IGF-I with high affinity) were present in all thyroid cancer histotypes. Because of recent evidence that isoform A of IR (IR-A) is a physiological receptor for IGF-II in fetal life, the relative abundance of IR-A in thyroid cancer was measured. Preliminary data indicate that overexpressed IRs mainly occur as IR-A in thyroid cancer. These data indicate that both IR/IGF-I-R hybrids and IR-A play an important role in the overactivation of the IGF system in thyroid cancer and in IGF-I mitogenic signalling in these tumours. J Clin Pathol: Mol Pathol
- insulin-like growth factor system
- insulin receptor
- insulin-like growth factor I receptor
Statistics from Altmetric.com
Pituitary thyroid stimulatory hormone (TSH) not only directly stimulates thyroid function but also exerts a trophic effect on the gland, as indicated by both its goitrogenic action and its ability to stimulate thyroid cancer proliferation.1 Therefore, L-thyroxine is given at TSH suppressive doses to patients who undergo surgery for differentiated thyroid cancer to prevent or reduce cancer growth.
Approximately 90% of thyroid carcinomas originating from the follicular epithelium have a papillary or follicular histotype. Although most are well differentiated and express TSH receptors, a proportion has a variable degree of dedifferentiation.2 Moreover, approximately 10% of all carcinomas are highly undifferentiated tumours that have completely or almost completely lost TSH responsiveness. Cancer mortality ranges from approximately 10% in differentiated tumours up to 100% in anaplastic tumours.1
Although TSH is the major growth factor for differentiated thyroid cancer, TSH independent cancer growth and metastatic spread is increasingly important with the progressive loss of differentiation. Therefore, TSH suppression with L-thyroxine administration is of limited efficacy in poorly differentiated or undifferentiated tumours.
A better understanding of the role of growth factors other than TSH might provide important insights into the biological mechanisms that affect thyroid cancer progression and may be useful for the design of new therapeutic approaches for dedifferentiated tumours.
It has been reported that a variety of tyrosine kinase growth factor receptors (including insulin-like growth factor (IGF-I), epidermal growth factor (EGF), Erb-B2, and hepatocyte growth factor (HGF)/scatter factor (SF) receptors) might be abnormally expressed and influence the biological behaviour of thyroid cancer.3–6
Early studies carried out in thyroid cell cultures have shown that the mitogenic effects of TSH are small in the absence of other growth factors, although they are greatly potentiated by the presence of insulin or IGF-I at physiological concentrations.7, 8 Using immunohistochemistry and in situ hybridisation, IGF-I and its receptor (IGF-I-R) have been shown to be expressed in thyroid cancer, suggesting that in these tumours the IGF-I-R is activated in an autocrine/paracrine manner.9, 10
IGF-I-R shares a high degree of homology with the insulin receptor (IR). Although IR is believed to be involved in metabolic regulation, recent work has suggested that IR might have a role in human cancer, including breast cancer.11 Both IR and IGF-I-R are tetrameric glycoproteins composed of two extracellular ligand binding domains (α subunits) connected by disulphide bonds to two cytoplasmic domains with tyrosine kinase activity (β subunits). The tyrosine kinase activity of these receptors (which is ligand stimulated) regulates major mechanisms that control cell growth and survival.12, 13
We will now briefly review novel aspects regarding the overactivation of the IGF system in thyroid cancer.
Insulin receptor overexpression: a key factor for the overactivation of the IGF system in thyroid cancer
To evaluate the involvement of the IGF system in thyroid cancer we measured IGFs and cognate receptors in both thyroid cell lines and tissue specimens.
First, we measured IGF-I content in extracts of tissue specimens from thyroid cancer and from normal thyroid. IGF-I content ranged from 104 to 2566.5 nM/g in cancer tissue and 69 to 680 nM/g in normal thyroid tissue. IGF-I mRNA, however, was not detected in a panel of eight thyroid cancer cell lines, whereas it was always present in cultured fibroblasts obtained from thyroid cancer specimens, indicating that stromal cells are the site of IGF-I production in thyroid cancer.14
Preliminary work indicates that IGF-II is locally produced in thyroid cancer. However, in contrast to IGF-I, it might be produced by malignant thyrocytes which, in contrast to normal thyrocytes can produce IGF-II (V Vella et al, 2001, unpublished data).
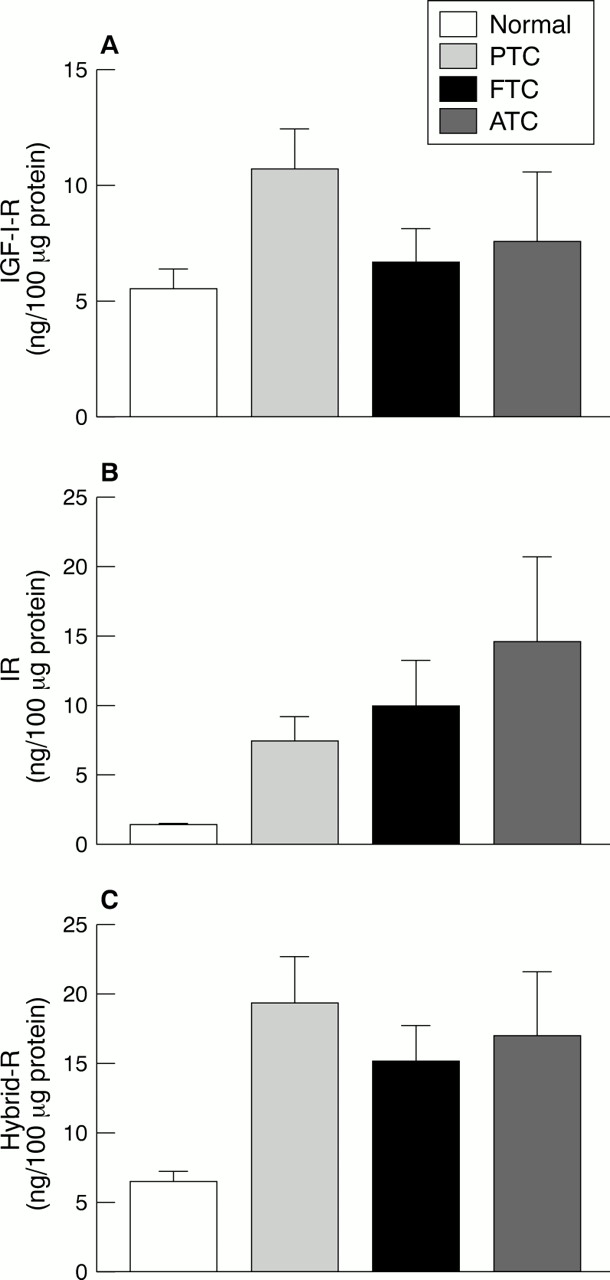
IGF-I-R content was greatly increased in thyroid cancer cell lines compared with normal primary cultures (range, 2.4 to seven times greater). However, mean IGF-I-R content was increased in well differentiated papillary carcinomas (approximately twofold higher than in normal tissue specimens) but not in poorly differentiated or undifferentiated cancer tissue specimens (fig 1A).14

(A) Insulin-like growth factor I receptor (IGF-I-R), (B) insulin receptor (IR), and (C) hybrid IR/IGF-I receptor (Hybrid-R) content (mean ±SEM) of normal thyroid (n = 7) and cancer thyroid specimens. PTC, papillary thyroid cancer (n = 8); FTC, follicular (poorly differentiated) thyroid cancer (n = 6); ATC, anaplastic thyroid cancer (n = 4).
In contrast, the median IR content was greatly increased not only in cancer cell lines (five to 28 times greater than normal thyrocytes) but also in thyroid cancer tissue specimens, with a trend towards higher values in less differentiated and anaplastic cancers (five to 7.4-fold increase) compared with well differentiated papillary cancers (fourfold increase) (fig 1B).
Interestingly, IR content was also increased in hypofunctioning benign thyroid adenomas, which have lost a differentiated function such as iodine uptake, but not in highly differentiated hyperfunctioning adenomas, indicating that IR overexpression is an early event in thyroid tumorigenesis.15
IR overexpression leads to increased formation of hybrid insulin/IGF-I receptors and increased IGF-I response
Hybrid insulin/IGF-I receptors (Hybrid-Rs) are heterodimers formed by one IR α and β subunit complex and one IGF-I-R α and β subunit complex. These hybrid receptors, formerly described in transfected cells and in the placenta, have since been found in most cells and tissues coexpressing IRs and IGF-I-Rs,16–18 and are believed to result from the random assembly of IR and IGF-I-R emireceptors, given their high degree of homology. Hybrid-Rs appear to bind with high affinity to IGF-I but not insulin.18 Therefore, we evaluated the possibility that Hybrid-Rs play an important role in mediating the effects of IGF-I in thyroid cancer.
Until recently, studies focusing on the role of Hybrid-Rs as compared with IRs and IGF-I-Rs have been difficult to perform because of the high structural and functional homology between these receptors. In particular, no direct assay was available for the direct measurement of Hybrid-Rs, and the proportion of these receptors was evaluated by the proportion of bound 125I-IGF-I that was immunoprecipitated by anti-IR antibodies.
However, using a recently developed specific enzyme linked immunosorbent assay, which allowed us to measure Hybrid-R content directly,19 we found that large quantities of Hybrid-Rs are expressed in thyroid cancer.14 Hybrid-Rs were increased (median values three to four times higher than normal) not only in tissue specimens from well differentiated papillary carcinomas (as were IGF-I-Rs), but also in specimens from poorly differentiated and undifferentiated carcinomas, probably as a consequence of increased IR expression (fig 1C).
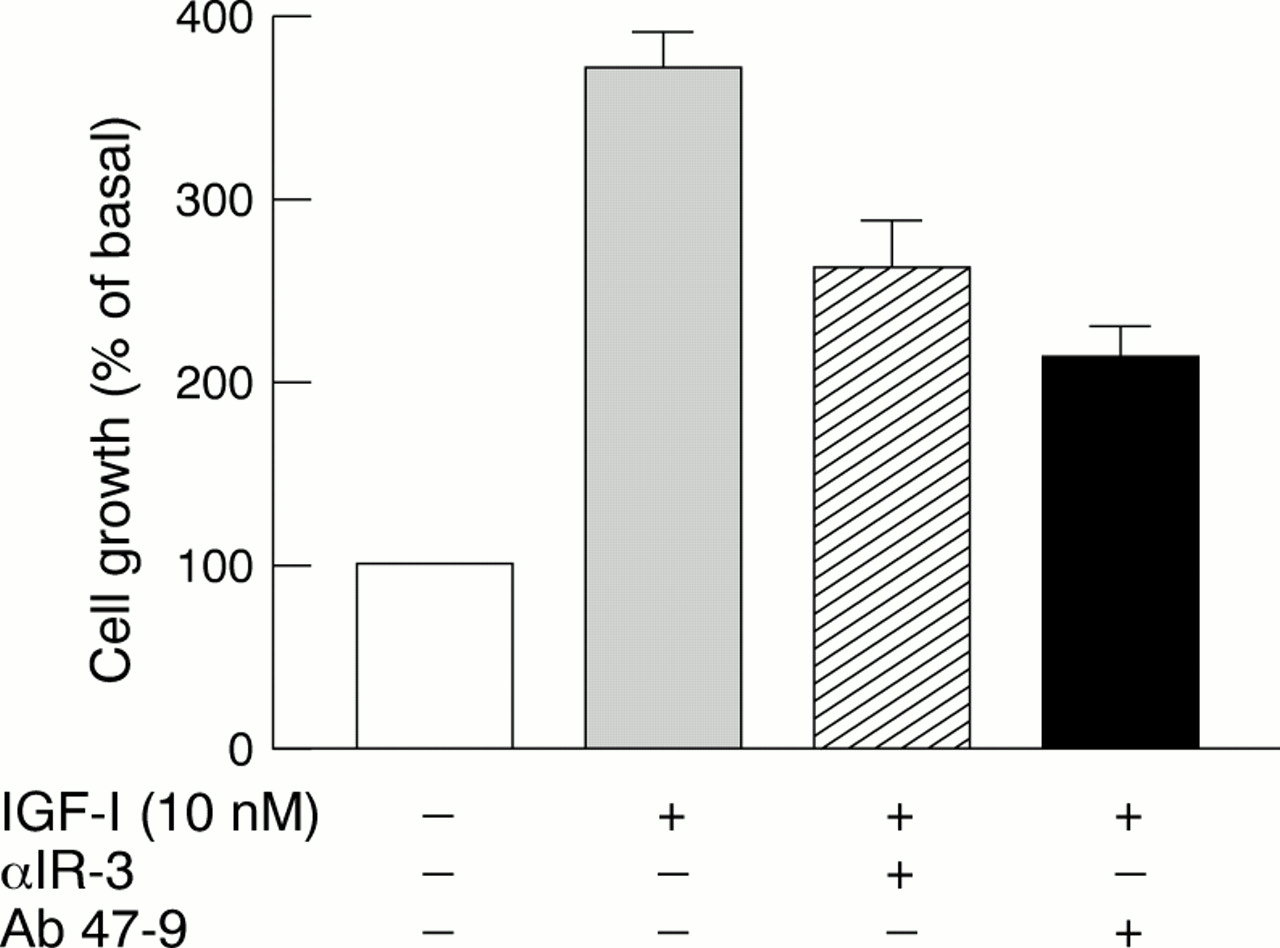
Hybrid-Rs accounted for 52–75% of the total IGF-I binding sites and their values were in close agreement with those expected for random assembly from IR and IGF-I-R half receptors. Individual tumours and cell lines could be identified with a high (2–4 : 1) Hybrid-R : IGF-I-R ratio. In these cells, Hybrid-R activation accounted for at least 50% of the effects of IGF-I, as evaluated by both receptor autophosphorylation and cell proliferation studies. In particular, in BC-PAP papillary cancer cells, Hybrid-R blockade with Ab 47-9 (a monoclonal antibody that does not recognise typical IGF-I-Rs) inhibited the growth response to IGF-I to a similar extent to that seen with IGF-I-R blockade by antibody αIR-3 (fig 2).14
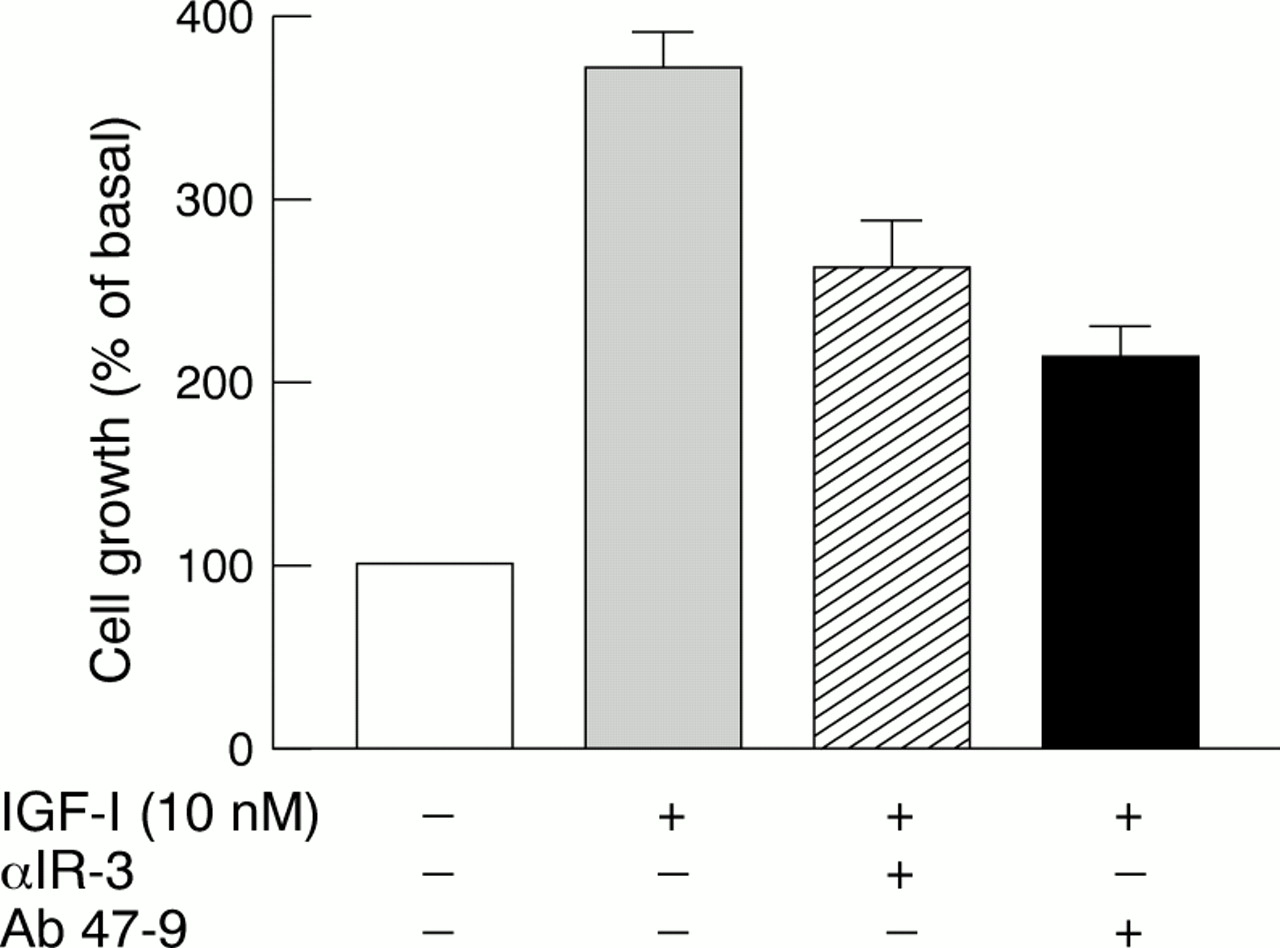
Thyroid cancer BC-PAP cell growth stimulated by insulin-like growth factor I (IGF-I) in the absence or the presence of antibodies blocking either IGF-I receptors (αIR-3) or hybrid insulin/IGF-I receptors (Ab 47-9).
IR-A overexpression may directly mediate IGF-II effects
Our recent studies on IR isoforms have provided new evidence of an additional role of IR overexpression in the activation of the IGF system in thyroid cancer.
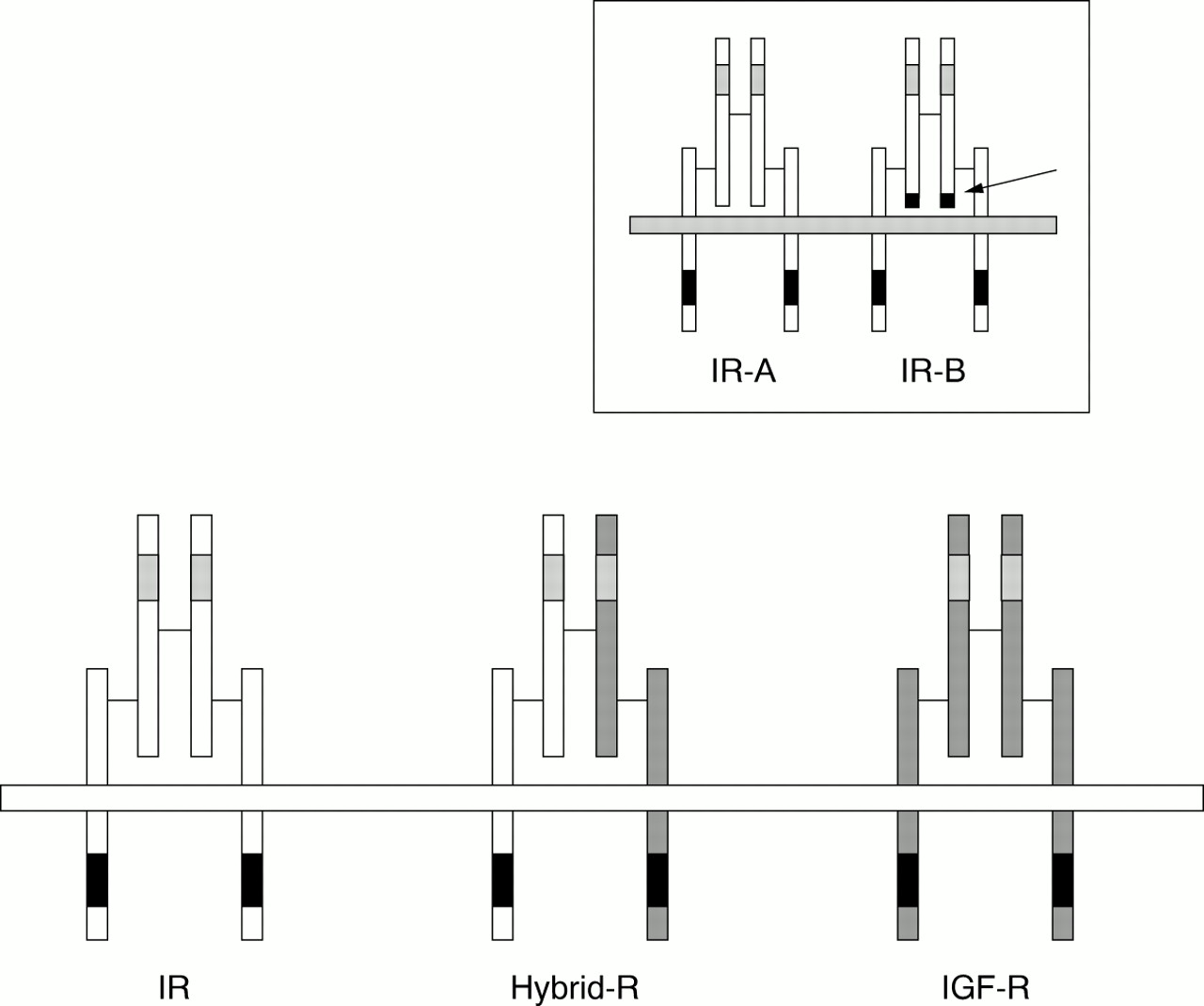
The human IR is present in two isoforms resulting, from alternative splicing of a small exon (exon 11) of the insulin receptor gene. The IR-A isoform does not contain 12 amino acids encoded by exon 11 at the C-terminus of the receptor α subunit, whereas the IR-B isoform does (fig 3).20 The relative expression of the two isoforms is tightly regulated in a tissue specific manner,20 although only small functional differences had been observed, with IR-A having a slightly higher binding affinity and IR-B having a more efficient tyrosine phosphorylation activity.

{kind=link}
{kind=link}
{kind=link}
Structural diagrams of the insulin receptor (IR), the insulin-like growth factor I receptor (IGF-I-R), and the hybrid IR/IGF-I receptor (Hybrid-R). Insert: structural schematics of the two IR isoforms (arrow indicates the location of the 12 amino acid residues encoded by exon 11 at the C-terminus of the α subunit).
We found that the IR-A isoform binds both insulin and IGF-II with high affinity.21 To avoid the interference of IGF-I-R, we used R− cells (engineered mouse embryo fibroblasts that do not express the IGF-I-R) transfected with either IR-A or IR-B cDNAs. In these cells, IGF-II bound to IR-A with an affinity similar to the binding affinity of IGF-I-R, and elicited a mitogenic effect quantitatively similar or even more pronounced than after exposure to insulin. The increased mitogenic potency of IGF-II compared with insulin is possibly related to different activation kinetics of receptor and postreceptor intracellular signalling molecules.21
In agreement with our findings, previous studies have shown that the activation of IR may elicit mitogenic and antiapototic effects similar to the activation of the closely related IGF-I-R, and that IGF-II may activate IR in certain cell models.22–27
The IR-A isoform is found predominantly in the fetus and the IR-B isoform in the adult. Interestingly, IR-A has also been found to be overexpressed in three major human tumours: breast, lung, and colon cancer.21 In breast cancer IGF-II, secreted in a paracrine manner, was a potent IR-A activator and promoted growth via this receptor.28
Our preliminary work in thyroid cancer indicates that overexpressed IRs predominantly occur as isoform A and that malignant thyrocytes acquire the ability to secrete IGF-II (V Vella, 2001, unpublished data). Thus, malignant transformation activates a newly identified autocrine loop in thyroid cells, involving IGF-II and the IR-A.
Conclusion
Different strategies have been proposed and implemented to inhibit cancer growth by blocking IGF-I-R binding and function. However, blockade of the IGF-I receptor does not completely prevent IGF-I stimulated growth,29–31 raising the possibility that the signalling system for IGF-I is complex and that other receptors are involved.
In thyroid cancer, as well as in other tumours, IGF-I-R receptors are overexpressed and both IGF-I and IGF-II are produced locally, in either a paracrine or autocrine manner. In addition, the concomitant overexpression of IRs adds complexity to this picture. First, some IRs form hybrids with IGF-I-Rs, thus expanding the pool of IGF-I binding sites. Second, IRs may directly bind IGF-II with high affinity because in many tumours they are predominantly present as the fetal IR-A isoform (fig 3).
A better understanding of these new aspects of IGF system overactivation in thyroid cancer may be relevant to therapeutic strategies aiming to block the effects of IGF-I, especially in dedifferentiated and metastatic tumours that are not responsive to TSH suppression with L-thyroxine.
Acknowledgments
This work was supported in part by the Associazione Italiana per la Ricerca sul Cancro (AIRC), MURST Cofin99 (to AB), and MURST 98 progetto biomedicina V8.