Article Text

Abstract
Objectives A functional polymorphism leading to reduced levels of growth and differentiation factor 5 (GDF5) was recently identified as a susceptibility factor for osteoarthritis. The authors studied the potential mechanisms of GDF5 involvement in osteoarthritis using haploinsufficient Gdf5Bp-J/+ mice.
Methods Gdf5Bp-J/+ mice were challenged in the collagenase-induced arthritis model, the medial meniscus destabilisation model, the papain-induced arthritis model and a treadmill running model. Bone density and subchondral bone parameters were determined using dual energy x-ray absorptiometry and peripheral quantitative CT. Additional in-vitro and ex-vivo analyses studied cartilage metabolism, gait and collagen characteristics.
Results Gdf5Bp-J/+ mice appeared phenotypically normal. No difference in osteoarthritis severity was found in the different models, with the exception of increased synovial hyperplasia in the joints of Gdf5Bp-J/+ mice in the treadmill model. However, in the collagenase-induced model severe joint damage was found in the contralateral joints of Gdf5Bp-J/+ mice. Gait analysis demonstrated an aberrant walking pattern in Gdf5Bp-J/+ mice. In addition, Gdf5Bp-J/+ mice have a decreased subchondral bone density and a distorted arrangement of collagen fibres in bone.
Conclusions These data suggest that decreased GDF5 levels in mice can contribute to osteoarthritis development by different mechanisms including altered loading and subchondral bone changes. This highlights the importance of the joint as an organ with different tissues involved in joint disease.
Statistics from Altmetric.com
A single nucleotide polymorphism (SNP) in the 5′ untranslated region of the human growth and differentiation factor 5 (GDF5) gene is associated with knee and hip osteoarthritis.1,–,3 The SNP affects allelic expression of the gene with the susceptibility variant leading to reduced levels of GDF5 in different joint tissues.4 Osteoarthritis is the most common form of chronic joint disease, with a significant impact on quality of life and a high socioeconomic burden.5 Osteoarthritis is characterised by pain and progressive loss of joint function, with gradual deterioration of the articular cartilage and mild inflammation of the synovium. This is accompanied by sclerosis of the subchondral bone and formation of osteophytes, bony outgrowths at the joint margins, probably in an attempt to restore joint stability. Osteoarthritis of the knees and hips is a complex disorder in which both genetic and acquired factors interact. Risk factors include obesity, joint malalignment, overuse and trauma.6
GDF5, also known as cartilage-derived morphogenetic protein 1, is a member of the transforming growth factor beta–bone morphogenetic protein family.7 GDF5, GDF6 and GDF7 are grouped within the GDF subfamily based on the high degree of homology in their mature signalling domain. The gene was discovered in the brachypodism (bp) mouse, in which a spontaneous mutation leads to the absence of proximal interphalangeal joints, fusions of bones in the wrist and ankle and abnormalities in the shape of bones associated with joint dislocations.8 Sequencing revealed a causal missense mutation in the GDF5 gene, resulting in complete loss of GDF5 function. Similar to observations in the bp mouse, several skeletal malformation syndromes occur in humans due to the loss of GDF5 function, such as acromesomelic chondrodysplasia Hunter–Thompson type,9 chondrodysplasia Grebe type,10 both autosomal recessive disorders, and brachydactyly type C,11 an autosomal dominant disorder. The different defects are characterised by a shortened appendicular skeleton with missing joints and fused bones.
Gdf5 is expressed in the condensing mesenchyme and in the joint interzone, the area where the future joint will form12 13 during development, and plays a role in appendicular joint formation12,–,14 and in the development of tendons and ligaments.15,–,17 Low levels of GDF5 are also found postnatally in different joint structures such as the articular cartilage, bone, synovium, tendons, ligaments and menisci. A role for GDF5 in the healing of bones, tendons and ligaments after trauma has been proposed.18,–,22 GDF5 is found in normal and osteoarthritis cartilage and in-vitro experiments have demonstrated that GDF5 increases extracellular matrix synthesis.20 23
These observations suggest that GDF5 plays both a role in musculoskeletal development and in postnatal joint homeostasis. As GDF5 appears to be present in most tissues of the adult joint, we and others anticipate that its role in osteoarthritis may not be restricted to one tissue.4 Homozygous Gdf5Bp-J/Bp-J mice have severe developmental defects such as joint fusions and dislocations precluding their analysis in osteoarthritis models. We therefore studied the effect of Gdf5 hypomorphism in Gdf5Bp-J/+ on the development of osteoarthritis in mice.
Methods
Mice
The Gdf5-Bp-J allele is the result of a spontaneous mutation in A/J mice (Jackson Laboratories, Bar Harbor, Minnesota, USA).8 Breeding strategy, genotyping and phenotype analysis, detailed description of the models and statistical analysis are found in supplementary materials, available online only.
Gait analysis
Footprint analysis was performed as described.24 25 The animal's hind paws were inked and footprints were made while the animal walked along a 1-m long and 7-cm wide runway with a paper-covered floor. Each mouse had to run three times. Every successful run (sequences of at least four consecutive steps) by 13 mice per group was taken into account for data analysis. The base of support was determined as the distance between the central pads of the hind paws. Stride length was determined as the distance between the central pads of two consecutive footprints at the same side.
Bone analysis
Total body (subcapital) bone density, lean and fat body mass of 8-week-old mice were determined by dual energy x-ray absorptiometry using a Piximus densitometer (Lunar, Madison, Wisconsin, USA). Trabecular, cortical and subchondral bone mineral content (BMC) of 8-week-old mice were assessed by peripheral quantitative CT (pQCT) using an XCT Research M+ system (Norland Medical Systems, Trumbull, Connecticut, USA). For trabecular and cortical BMC, slices of 0.2 mm in thickness were scanned using a voxel size of 0.07 mm. Three scans were taken 2.4–2.6 mm from the distal end of the femur or 1.4–1.6 mm from the proximal end of the tibia to determine trabecular bone parameters. An additional scan was taken 4 mm from the distal end of the femur or 7 mm from the proximal end of the tibia to determine cortical bone parameters. For subchondral bone measurements, slices of 0.1 mm in thickness were scanned using a voxel size of 0.07 mm. Three scans were taken from the extreme distal end of the femur after manual controlled positioning.26
Results
Characterisation of Gdf5Bp-J mice
Gdf5Bp-J/Bp-J mice carry a mutation causing a premature translational stop codon in the Gdf5 gene leading to loss of function.8 Gdf5Bp-J/+ heterozygous mice have no apparent skeletal abnormalities and show normal growth in contrast to Gdf5Bp-J/Bp-J mice (supplementary figure 1, available online only). No obvious knee abnormalities were observed in Gdf5Bp-J/+ mice compared with wild-type (WT) mice by radiographic (supplementary figure 1, available online only) and histological analysis (data not shown). Gdf5Bp-J/+ mice showed identical weight curves compared with their control littermates, whereas the weight curves of Gdf5Bp-J/Bp-J mice were approximately 20% lower (supplementary figure 1, available online only). Also the length of the long bones was comparable between heterozygous and WT mice (supplementary figure 1, available online only), whereas in Gdf5Bp-J/Bp-J mice the bones were approximately 20% shorter (data not shown). Subcapital total body measurements with dual energy x-ray absorptiometry at the age of 8 weeks showed no difference in bone mineral density (BMD), BMC, bone area, lean and fat mass between both groups. More specific densitometry analysis with pQCT demonstrated a significantly decreased trabecular area in the long bones of Gdf5Bp-J/+ mice at the age of 8 weeks (femur 1.54±0.12; tibia 1.30±0.19) compared with WT mice (femur 1.66±0.13, p≤0.027; tibia 1.50±0.17, p≤0.014) thereby confirming haploinsufficiency in heterozygous mice. No differences in cortical bone parameters were found (data not shown).
Effect of Gdf5 haploinsufficiency on development of osteoarthritis in mice
We studied the effect of Gdf5 haploinsufficiency on osteoarthritis in four different mouse models in Gdf5Bp-J/+ mice and WT littermates. Each of these models highlights specific factors that contribute to the disease in humans. In healthy 8-week-old mice, the sulphated proteoglycan content of the articular cartilage was not different between heterozygous and WT mice (data not shown). Twenty-one days after the induction of collagenase-induced osteoarthritis in Gdf5Bp-J/+ mice and WT littermates, typical histological changes were seen in the collagenase-treated knees (proteoglycan loss, osteophyte formation and synovitis; figure 1). Collagenase treatment resulted in an increased sulphated proteoglycan loss in WT and heterozygous articular cartilage (figure 2A). No difference in cartilage damage was seen between WT and Gdf5Bp-J/+ mice in the collagenase injected knees. Loss of sulphated proteoglycans in phosphate-buffered saline (PBS)-treated control knees was significantly increased in GDF5Bp-J/+ mice compared with WT mice (p<0.036), suggesting that the instability and progressive damage of the right knee also affected joint homeostasis in the contralateral knee (figure 1 and figure 2A). In collagenase-injected knees, scores for synovial hyperplasia, infiltration and capsule fibrosis, a feature typical of mouse models of osteoarthritis (figure 2B) were comparable between heterozygous and WT mice. In the contralateral (PBS injected) knees, however, scores for synovial hyperplasia and capsule fibrosis were significantly increased in the heterozygous group compared with the WT group (p<0.044 and p<0.039, respectively) (figure 2B). Moreover, a comparable amount of WT (8/9) and heterozygous (7/9) mice had osteophytes formed in the collagenase-injected knees, but heterozygous mice (4/9) showed more osteophytes in the contralateral knees compared with WT mice (0/9).

Collagenase-induced osteoarthritis in Gdf5Bp-J/+ heterozygous (HZ) and Gdf5+/+ wild-type (WT) mice: representative safranin O-stained sections of WT and heterozygous, collagenase (right) or phosphate-buffered saline (PBS; left)-treated knees. Arrows indicate infiltration and fibrosis, asterisk indicates osteophytes.

(A) Digital image analysis of articular cartilage in collagenase and PBS injected knees from heterozygous (HZ; black bars) and wild-type (WT; grey bars) mice (N=7 and 9, 9 and 11 per group respectively; data presented as mean±SEM; Kruskal–Wallis test followed by Mann–Whitney U test: overall significance p<0.001; 1/1 *p<0.006, **p<0.001). (B) Individual pathological scores for synovial hyperplasia, infiltration and fibrosis 21 days after injection with collagenase or phosphate-buffered saline (PBS) in heterozygous and WT mice. Score ranging from absent (=0) to mild (=1), moderate (=2) and severe (=3) (N=9 per group; data presented as individual score per mouse (diamond)+mean score (dashed line); Kruskal–Wallis test followed by Mann–Whitney U test: overall significance for synovial hyperplasia p<0.002, infiltration p<0.0001 and fibrosis p<0.002; 1/1 *p<0.044, **p<0.039, (*)p<0.052).
The collagenase model is a model of severe instability. The contralateral changes suggest that Gdf5Bp-J/+ mice are more prone to develop osteoarthritis possibly secondary to increased load of the unchallenged knee. In a pilot experiment we studied signs of osteoarthritis in 26-week-old mice (five Gdf5Bp-J/+ and three WT mice). In three of 10 knees from Gdf5Bp-J/+ mice discrete osteophytes were found, as was the case in one of six knees from WT mice. There was no significant difference in synovial hyperplasia, infiltration or fibrosis between the groups. Digital image analysis of the articular cartilage revealed no differences in cartilage damage scores between Gdf5Bp-J/+ and WT mice (data not shown). The absence of a clear spontaneous phenotype at 26 weeks stimulated us to consider another approach. We therefore studied the development of osteoarthritis-like changes by forced treadmill running during 3 weeks (treadmill model) in 16-week-old Gdf5Bp-J/+ mice and WT mice. Mechanically induced osteoarthritis is characterised by articular cartilage damage, formation of osteophytes and mild synovial hyperplasia and capsule fibrosis. We did not find significant differences in cartilage damage in Gdf5Bp-J/+ mice compared with WT (cartilage damage index 1.55±0.03 vs 1.57±0.03 and 1.68±0.05 vs 1.71±0.05 in the tibia and femur, respectively (heterozygous vs WT) p>0.05). However, a significantly increased score for synovial hyperplasia was found in the heterozygous group compared with the WT group (p<0.03) and the capsule fibrosis scores were also increased; however, they were not significantly different (p<0.08; figure 3A). In contrast, no differences in cartilage damage, synovial hyperplasia, infiltration and capsule fibrosis were found between Gdf5Bp-J/+ mice and WT mice in the meniscus destabilisation model (data not shown) and in papain-induced arthritis (data not shown). Also, in-vitro cartilage breakdown assays did not show differences between Gdf5Bp-J/+ and WT mice (data not shown). Histological analysis (H&E, safranin O, Von Kossa, toluidine blue and alizarin red S) of hip cartilage of WT and Gdf5Bp-J/+ mice showed no apparent differences (data not shown), suggesting that factors other than impaired cartilage homeostasis may cause the increased susceptibility to osteoarthritis in heterozygous mice.

(A) Forced treadmill running in Gdf5Bp-j/+ heterozygous (HZ) and Gdf5+/+ wild-type (WT) mice: individual pathological scores for synovial hyperplasia, infiltration and fibrosis in knees of heterozygous and WT mice after forced treadmill running. Score ranging from absent (=0) to mild (=1), moderate (=2) and severe (=3) (N=8 mice per group, two knees per mouse; data are presented as individual score per mouse (diamond)+mean score (dashed line); Mann–Whitney U test *p<0.03, (*)p<0.08). (B) Gait, bone and collagen type I analysis of Gdf5Bp-j/+ (HZ) and Gdf5+/+ (WT) mice. Mice with inked hind paws had to run over a 1-m long, 7-cm width runway, covered with paper. (C) Distance was measured between the central pads of two consecutive footprints at the same side (N=60 and 56 successful runs of 13 mice (per group) that were each allowed to try a run three times; data are presented as median, quartiles and percentiles, Student's t test *p<0.05).
Gait analysis of Gdf5Bp-J/+ mice
We therefore hypothesised that altered joint loading and instability may be a factor in the development of osteoarthritis in the contralateral knee in the collagenase model. Gait analysis of healthy 8-week-old Gdf5Bp-J/+ and WT mice demonstrated a significant decrease in stride length (distance between two consecutive steps; figure 3 B,C) in Gdf5Bp-J/+ mice compared with WT mice (p<0.05), while the base of support distance (distance between left and right foot) did not change (data not shown). These data show that Gdf5Bp-J/+ mice have an aberrant walking pattern suggesting a decreased joint stability changing the mechanical load in the joint. When severe instability is present in the index joint in the collagenase-induced osteoarthritis model increased load on a less stable left knee may be the trigger for osteoarthritis development.
Bone–subchondral bone analysis
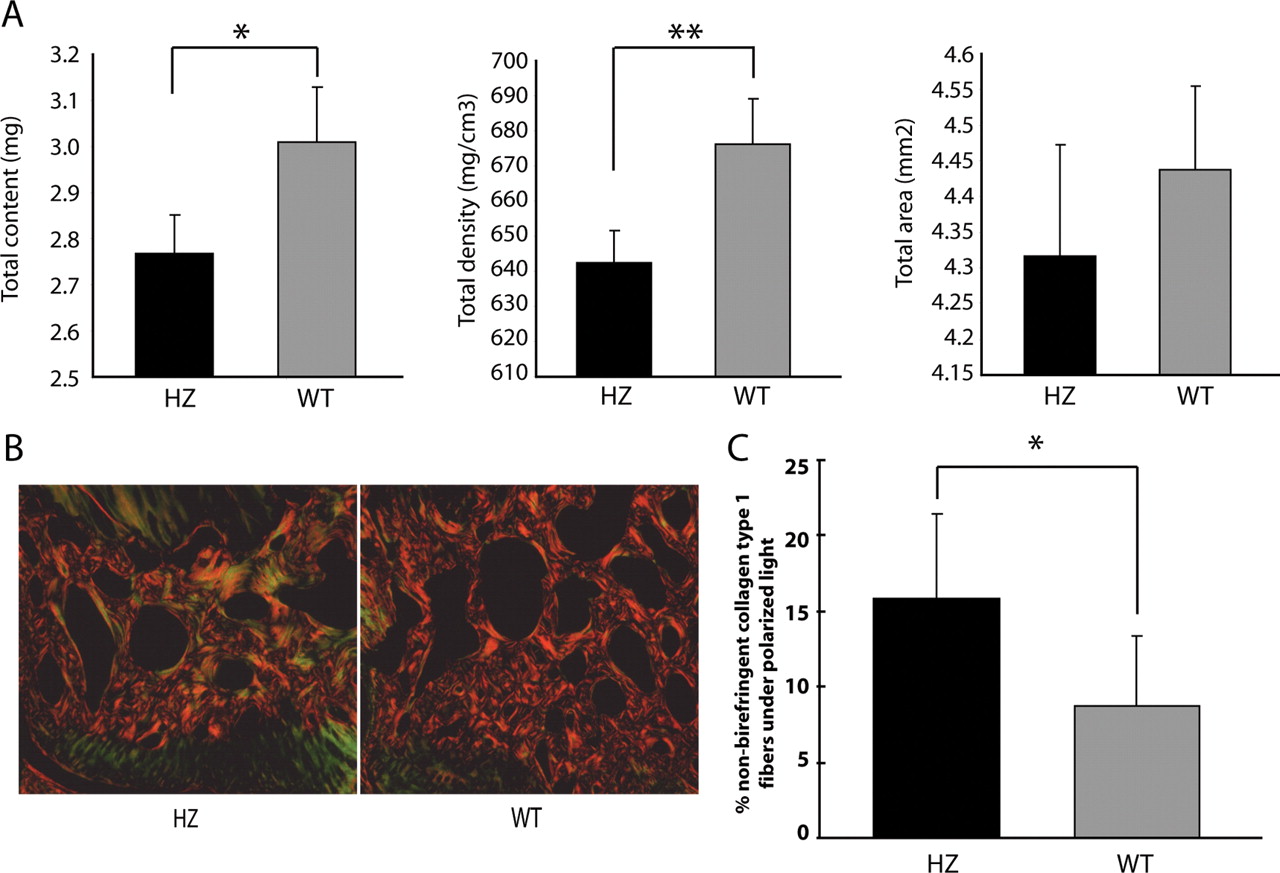
We also performed specific densitometry analysis with pQCT of the subchondral bone (figure 4A), demonstrating a significantly diminished subchondral bone content and density (p<0.05) in Gdf5Bp-J/+ mice compared with WT mice at the age of 8 weeks. Moreover, when we checked the orientation of collagen fibres in the subchondral bone area of picrosirius red-stained knee joints with the aid of polarisation microscopy (figure 4B), we found more non-birefringent collagen fibres (green) representing less organised collagen in the subchondral bone of Gdf5Bp-J/+ mice compared with WT mice (yellow-red; p<0.007; figure 4C). Analysis of the fibres in the cartilage and ligaments did not suggest differences between the Gdf5Bp-J/+ and WT mice. However, abnormal organisation and orientation cannot be excluded in these tissues as fibres in the cartilage are more difficult to distinguish under polarised light, whereas the fibrous ligaments are not confined to the conventional cutting planes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Peripheral quantitative CT measurements of the subchondral bone for total bone content, density and area (N=11 mice per group, data presented as mean±SEM; Student's t test *p<0.003, **p<0.0001). (B) Representative images of picrosirius red-stained sections of heterozygous (HZ) and wild-type (WT) under polarised light. Arrows indicate birefringent collagen type I fibres (red-yellow), arrowheads indicate non-birefringent collagen type I fibres (green). (C) Percentage of non-birefringent collagen type I fibres, measured by Image J, in subchondral bone area of heterozygous (black bars) and WT (grey bars) mice (N=5 mice per group, two compartments per mouse (medial and lateral tibial subchondral bone); data are presented as mean±SD; Student's t test *p<0.007).
Discussion
In this study, we demonstrated that Gdf5Bp-J/+ mice show increased osteoarthritis-like changes in the contralateral knees in the collagenase model and a higher degree of synovial hyperplasia in a running model. These findings are associated with an altered gait pattern in heterozygous mice and with changes in the subchondral bone and collagen fibres. In accordance with a recent study on the effect of the GDF5 osteoarthritis susceptibility allele in different joint tissues,4 these data support a role for GDF5 in the biology and pathology of the postnatal joint organ.
Loss of Gdf5 in bp mice not only results in a number of severe skeletal malformations,8 but also affects collagen type I-rich tissues such as tendons, ligaments, bone and skin.27 Homozygous bp mice failed to develop intra-articular ligaments,15 although the anatomy of heterozygous mice appears normal. Analysis of the Achilles tendon in Gdf5Bp-J/Bp-J mice demonstrated changes in its ultrastructure, mechanical properties and composition. These tendons are structurally weaker and more compliant with increased laxity.16 The human chondrodysplasias Grebe and Hunter–Thompson are also characterised by laxity of the joints with dislocations. Whether this is due to a direct effect of GDF5 on collagen fibrillogenesis or an indirect effect remains to be elucidated. We did not detect changes in fibre orientation or organisation in knee joint-associated ligaments. However, technical issues with processing and cutting precluded analysis of isolated joint-associated ligaments. The surprising results in the collagenase model with osteoarthritis-like changes on the contralateral control side in heterozygous mice and the altered gait pattern suggest that Gdf5 haploinsufficiency affects the stabilising capacities of joint connective tissues upon loading and thereby contributes to osteoarthritis. The more discrete changes in the treadmill running model are also in line with this hypothesis. In contrast, mild instability in the medial meniscus destabilisation model does not appear sufficient to cause differences between heterozygous and WT mice. This could be explained by either a lack of changes in loading in the control joint in the model or a lack of sensitivity of the scoring system used in the index joint. Our observations have their limitations as they cannot prove that instability is the determining factor because neuromuscular changes in the mutant mice may also be contributing to the abnormal gait.
The specific effect in the treadmill running model on synovial hyperplasia is intriguing. Synovitis is well recognised in osteoarthritis patients, and recent studies have suggested that a number of cytokines appear to be involved.28 Synovitis and joint pain in osteoarthritis also appear to be well correlated.29 In our experiment, we only noticed synovial hyperplasia without inflammatory cell infiltration. Synovial hyperplasia in the exercise model could be one of the earliest signs of joint damage or strain as the lining layer cells are sensitive to a number of factors including molecules released from the cartilage. This could occur before morphological changes become apparent under the microscope. The synovial hyperplasia may also be specifically related to the hypothesised joint instability in the mutant mice and a consequence of increased load.
Bone analysis revealed that Gdf5Bp-J/+ mice had a significantly lower trabecular area in their long bones, but no differences in cortical parameters. Moreover, the subchondral bone showed a decrease in BMC and BMD. The role of the bone–cartilage biomechanical unit and the eventual primacy of subchondral bone changes in the development of osteoarthritis is still a matter of debate. From a functional perspective any imbalance in the bone–cartilage unit either due to sclerotic subchondral bone or cortical bone26 leading to cartilage deformation upon movement against a higher resistance as well as soft subchondral bone leading to differences and less effective stress distribution can contribute to osteoarthritis. For instance, Brtl mice, having a genetic collagen type I defect, rapidly develop osteoarthritis and cartilage degradation, which is explained by an altered architecture of the underlying subchondral bone.30 Similar factors could play a role in the load and exercise osteoarthritis phenotype of Gdf5Bp-J/+ mice. Of interest, this has only been suggested up till now in structural molecules that are part of the extracellular matrix and not in growth factors. For instance, abnormal collagen fibrils in tendons, due to deficiency in biglycan, fibromodulin or lumican, small leucine-rich proteoglycans regulating matrix organisation in collagens fibrils, lead to gait impairment, joint laxity and spontaneous development of osteoarthritis.31 32
In this study, we did not demonstrate a direct effect of Gdf5 on the articular cartilage. Nevertheless, the expression of GDF5 in human healthy and osteoarthritic articular cartilage and its potency to stimulate proteoglycan matrix synthesis in vitro20 23 indicates its potential and likely role in cartilage homeostasis. This apparent discrepancy can be explained in different ways. The effect of Gdf5 haploinsufficiency in the articular cartilage in mice may be too discrete to have an effect on the osteoarthritis models used in this study. Also, increasing evidence suggest a functional redundancy of the closely related family members Gdf6 and Gdf7, which may help in maintaining cartilage homeostasis even when Gdf5 levels are reduced.33
In summary, the data presented here suggest that Gdf5 is involved in the biology and pathology of the whole joint organ and that its role reaches far beyond that of a cartilage metabolism regulating molecule. Finally, our biological data in the different models are supportive of the genetic data indicating an association between Gdf5 hypomorphism and osteoarthritis.1,–,3 More detailed studies in the human model should provide additional insights into the mechanisms relevant to the development and progression of osteoarthritis.
Acknowledgments
The authors would like to thank Professor Dr P Carmeliet for the use of the treadmill, Dr T Vincent and Dr J Inglis for teaching the surgical osteoarthritis model, Mrs J Peeters, Mrs A Hens, Mr E Van Herck and Mrs A Similon for their technical assistance with this study.
References
Supplementary materials
Web Only Data ard.2010.134619
Files in this Data Supplement:
{kind=link}
Footnotes
-
Funding This work is funded by a grant from Coordination Theme 1 (Health) of the European Community's FP7, grant agreement number HEALTH-F2-2008-00 (large collaborative project TREAT-OA) and by a GOA grant from Katholieke Universiteit Leuven, Leuven. MD is the recipient of a doctoral fellowship from the Institute for the Promotion of Innovation through Science and Technology in Flanders (IWT Vlaanderen). RJUL is the recipient of a postdoctoral fellowship from the Research Foundation, Flanders (FWO Vlaanderen).
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.